 Print
Print  PDF
PDF
Ритуализованная угроза больших пёстрых дятлов. Источник Светлана Новосёлова
Резюме. По завершению послегнездовых перемещений, в августе-сентябре, большие пёстрые дятлы занимают территории в плотных поселениях, и дальше охраняют их всё осень и зиму — крайне активно и жёстко. Именно в плотные поселения они стремятся в первую очередь, хотя там и занять, и удержать территорию гораздо труднее, и лишь потеряв её из-за неуспешной охраны, они «скатываются» в более разреженные поселения, т.
е. плотные выступают «регуляторами-распределителями» населения вида между поселениями разной плотности и устойчивости так, что в них концентрируются «лучшие» особи в плане территориальной агрессии. Обсуждается адаптивный смысл затрат владельцев на разные формы территориального поведения в течение всей осени и зимы, поскольку они а) довольно значительны и б) не «сокращаются» даже в периоды сильных морозов,. Показывается, что здесь неприложимы объяснения, связанные с немедленным выигрышем, т.е. монополизацией запасов семян, которыми птицы кормятся, или же «содержащего» их пространства.Все территории владельцев в плотных поселениях содержат заведомый избыток корма, при невозможности его обеспечить, из-за неурожая или равным образом из-за поражений в конфликтах, сокращающих охраняемое пространство, группировки просто не возникают (а территории оставляются птицами ещё до истощения шишки).
Проверена и подтвердилась гипотеза, что долговременная и жёсткая охрана территорий в плотных поселениях D.major преследует долговременный выигрыш в виде сильного сокращения период поиска партнёра и более ранних сроков образования пар весной, после нескольких месяцев постоянных попыток владельцев переделить в свою пользу охраняемое пространство группировки.
Последнее ведёт к социальной дифференциации — статусов, моделей поведения и внутренней структуры территорий владельцев, описанной в работе. И именно в плотных поселениях максимально сокращены начальные этапы образования пар, у больших пёстрых дятлов наиболее длительные и затруднённые из-за высокого риска срыва начавшегося ухаживания территориальной агрессией, т.к. сосед противоположного пола при подобных сближения воспринимается также как потенциальный захватчик. Показано уменьшение этого риска благодаря социальной дифференциации, происходящей именно в плотных поселениях на протяжении всей осени и зимы. Весной, в феврале-марте, сближение и попытки ухаживания птиц близких статусов вызывает гораздо меньше агрессии и позволяет гораздо быстрее образовать пару, чем в разреженных, где весеннее возбуждение началось в те же сроки. Иными словами, «выигрыш» от территориальной активности в течение осени и зимы реализуется только весной.Введение
Содержание
В августе-сентябре большие пёстрые дятлы занимают одиночные территории в плотных поселениях (сеть их формируется в одних и тех же местах обильно плодоносящих сосняков и ельников), и активно охраняют их до марта-апреля. С началом весеннего возбуждения (январь-февраль, реже в предновогодние оттепели декабря) эта территориальность сперва перестраивается, а потом прекращается. Продолжая охранять это пространство, владельцы (и владелицы) территорий одновременно сближаются с разными резидентами противоположного пола с ухаживательными демонстрациями, тестируя их «качество» как потенциальных партнёров1.
Образование пар: важность гарантий от срыва
Что довольно рискованно, особенно поначалу. Дятлы весьма агрессивны, особенно самки; резиденты всегда воспринимают «сближающихся» соседей как потенциальных захватчиков и могут атаковать прежде, чем эффекты ухаживания пересилят агрессию. Поначалу риск срыва ухаживания агрессией или уходом одной из птиц весьма велик во взаимодействиях со всеми соседями, с которыми этот дятел пробует сблизиться (40—60%). Далее во взаимодействиях с кем-то одним из них (или одной; у разных полов они полностью симметричны, отличаясь большей барабанной активностью у самцов и частотой криков «ре-ре-ре-ри2» у самок до или после слётов на чужую территорию) агрессия прекращается раньше всего, а сближение развивается особенно быстро.
Пара образуется там, где в подобных сближениях быстрее всего сокращается дистанция между самцом и самкой, токования повторяются чаще, длятся дольше, а срываются агрессией или уходом реже, сравнительно с взаимодействиями с другими соседями (-ками). Здесь же партнёры быстрей переходят к взаимной адресации всё более эффективных брачных демонстраций репертуара (рис.1А). Их действенность (рис.1Б) оценивали трояко
1) по отражению большего уровня сексуальной мотивации;
2) по эффективности стимуляции ещё большего сближения потенциальных партнёров в данном взаимодействии;
3) вероятности возобновления токования в случае срыва, оцениваемой частотой повторных сближений тех же птиц после срыва за время, меньшее среднего времени долбления шишки в данной группировке, т.е. по усилению последействия уже случившейся брачной коммуникации.
У разных видов птиц (практически всех, которых исследовали специально) скорейшее образование пар сравнительно со средней датой для популяции строго коррелирует с более ранним началом интенсивных копуляций партнёров и, соответственно яйцекладки. Т. е. с быстрейшим развитием ухаживания самца и самки на стадии консолидации пары и с репродуктивным успехом за сезон в целом. См. пример с розовой крачкой Sterna dougallii.
Оценки 1)—3) эффективности демонстраций, специфически связанных с контекстом ухаживания (их образы см. рис.1А) хорошо согласуются между собой (рис.1Б). Уровень мотивации в 1) независимо оценивали временем в днях, прошедшем от начала весеннего возбуждения именно в данной группировке. Для каждой из них было показано, что более поздно появляющиеся в репертуаре птиц демонстрации рис.1 более эффективны во всех трёх аспектах. Они стимулируют более тесное сближение потенциальных партнёров, до непосредственного телесного контакта включительно (№5—9), с ними связана большая вероятность возобновление токований после срыва – в сравнении с демонстрациями, появляющимися в репертуаре раньше (см. ось абсцисс рис.1). То же верно для «всей исследованной популяции», данные по которой получены усреднением наблюдений за всеми прослеживаемыми группировками во все сезоны. Здесь эта закономерность даже чётче.
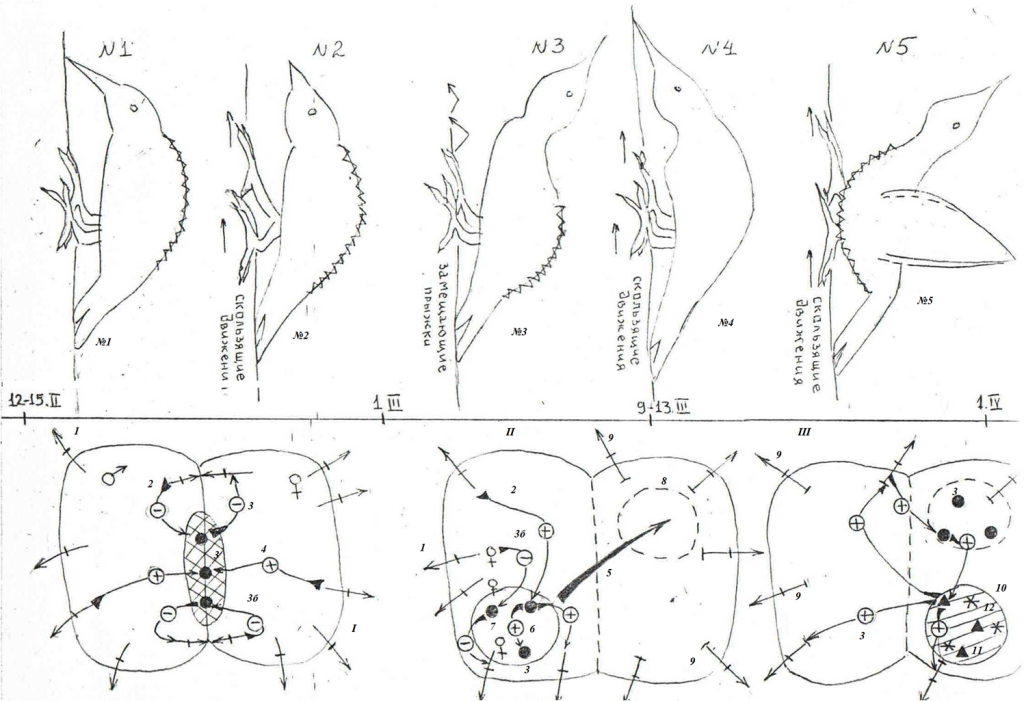
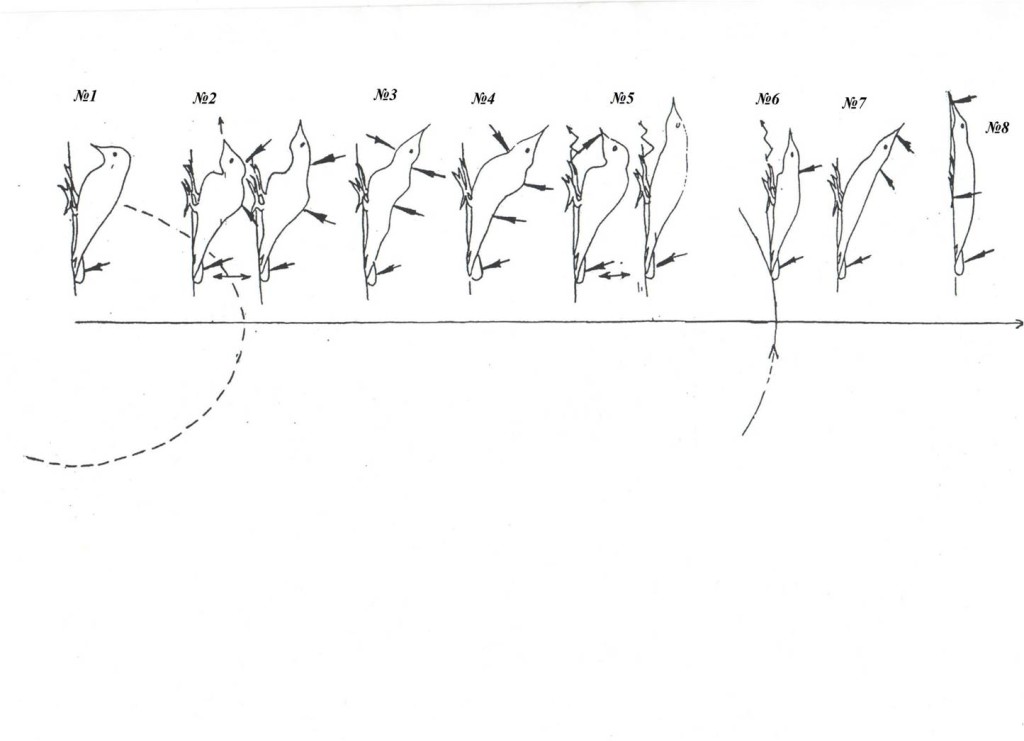
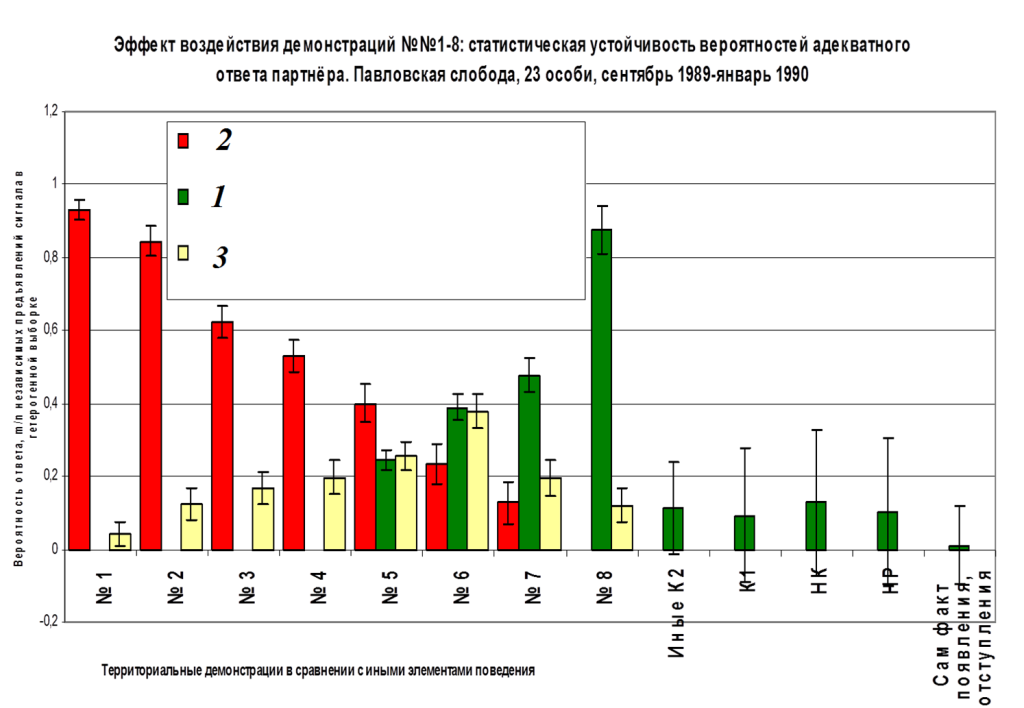
Рис. 1. Репертуар брачных демонстраций больших пёстрых дятлов (№№1-9, самцы и самки идентичны).
А. Выделенные демонстрации на мотивационном градиенте (абсцисса, время в днях от начала регулярной барабанной активности данной особи, абсцисса), схемы телодвижений. Ниже её — трансформация одиночных территорий потенциальных партнёров в парные по ходу ухаживания. Обозначения. Показаны территории самца и самки — будущих потенциальных партнёров, их охраняемые границы и функциональные зоны — начальных этапов тока у второй и поиска места для гнезда у первого.
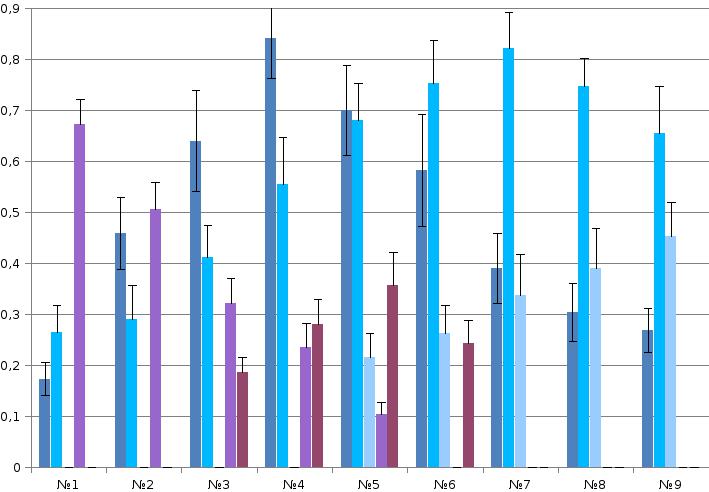
Рис. 1. Репертуар брачных демонстраций больших пёстрых дятлов (№№1-9, самцы и самки идентичны).
Б. Эффекты воздействия выделенных демонстраций №№1-9. Даны частоты реализации четырёх специфических двигательных ответов в выборке n независимых предъявлений: 1) ещё большего сближения с партнёром (сокращение дистанции, демонстрируя). 2) возобновления токования после срыва предыдущего за время, меньшее ⅔ времени долбления сосновой или еловой шишки, в зависимости от питания этой птицы. 3) Подставления (либо телесного контакта) и спаривания, 4) остановки сближения паузой, или срыва взаимодействия из-за агрессии (бегства) одной из птиц.
Примечание. Популярный рассказ о методах регистрации и анализа «развёрток» активности наблюдаемых особей во времени, позволяющий выделять единицы поведения (далее — «демонстраций») на чисто структурной основе, без неявного привнесения функциональных моментов, и затем оценивать их эффект, см. «Большая восьмёрка большого пёстрого дятла». См. также Фридман В.С. Коммуникация в агонистических взаимодействиях большого пестрого дятла // Бюлл. МОИП. — Сер. Биол. — 1993. — Т.98. — №4. — С.34–45; «От стимула к символу. Сигналы в коммуникации позвоночных животных», ч.2, разд. 6.3. «Методические проблемы расшифровки «языка животных»» (М.: URSS, 2013).
Наиболее эффективные демонстрации из разряда ухаживательных (самые «поздние» на рис.1А) появляются с начала—середины апреля. Тогда сближение достигнуто, самец с самкой преследуют друг друга и демонстрируют почти вплотную («клюв к клюву» или «хвост к носу», но не «бок о бок»). Пара активно подыскивает место для гнезда в соответствующей зоне внутри объединённой территории, на бывшем участке самца (рис.1А, 10). Дятлы закладывают серию инициальных дупел, своего рода «заготовок» гнездовых, и токуют у них (там же, 12). Позднее одно из них (где ток наиболее интенсивен и/или, прервавшись, чаще всего возобновляется сразу без перерыва на кормление), достраивается до гнездового.
Так или иначе, предъявление данных демонстраций, помимо возобновления токований, с определённой вероятностью вызывает подставление самки и/или покрытие её самцом (рис.1Б). Что происходит сперва с промедлением на 4—8 с, потом и немедленно (рис.1А, 11). «Сперва» — это конец марта в плотных поселениях, середина апреля в разреженных, «потом» — конец апреля-начало мая, когда гнездостроение в полном разгаре и/или завершено.
На любой выборке данных, локальной или объединённой, видим, что брачные демонстрации фиксируются в репертуаре дятлов тем позже, чем эффективней они в этом плане. Это верно для обоих их типов: и просто стимулирующих большее сближение птиц, продолжение либо возобновление тока после «срыва» (№№1-3), так и побуждающих к копуляции (№№6-9). Это «расставляет» на мотивационном градиенте все демонстрации, появляющиеся в связи с ростом весеннего возбуждения (рис.1А; «позже» означает «больше»), одновременно «ранжируя» пары по успешности образования в данном годовом цикле3. В «передовых» парах самец и самка раньше заканчивают сближения с соседями и дальше токуют только друг с другом, всё согласованней перемещаются по участку и пр. Ухаживание здесь развивается быстрей, раньше начинаются копуляции, в т.ч. на 5—12 дней раньше гнездостроения, раньше переход к яйцекладке и начало насиживания.
Увы, подобную дифференциацию «на всём протяжении» образования пар изучили только в разреженных поселениях (<10 особей/км2) Львовской области весной 1989 г., Аминьевского леса под Пущино весной 1998—2000 гг.4, в Пироговском лесопарке г. Мытищи весной 2002—2007 гг. В обсуждаемых здесь плотных поселениях это удалось лишь на части процесса.
В первом случае дятлы часто гнездятся в пределах осенне-зимних территорий или поблизости (до 1—1,5 км). Во втором — они исчезают на гнездовое время из данной местности, а птиц, гнездившихся несколько далее (в радиусе до 15 км) слишком мало для статистического анализа (19 за все годы). Это крайне удобно в исследованиях социальной стратификации во внегнездовых группировках, но позволяет судить о размножении их членов лишь по косвенным данным.
Так или иначе, после образования пары участки самца и самки объединяются, и только самец охраняет и рекламирует это пространство. В следующие 7—10 дней территориальность самок редуцируется до минимума5 (маркирование мест пребывания и, при нахождении поблизости, мест территориальных конфликтов самца). Из форм социальной активности у них остаётся только ухаживание и особенно «натравливание» своих самцов на соседних6, несколько интенсифицирующее охрану территорий на фоне её общего угасания в апреле. В это время основные силы самца чем дальше, тем больше уходят на гнездостроение. У самок оно же идёт на питание, нужное для образования большой кладки: 5—10 яиц у старых и 4—8 у молодых самок; изменчивость размера кладки см. Бутьев В.Т., Фридман В.С. Большой пёстрый дятел// Птицы России и сопредельных регионов. Совообразные—дятлообразные. М.: Т-во научн.изд. КМК, 2005.
Что значительно выше, чем у видов, специализированных к долблению ксилофагов, вроде белоспинного и трёхпалого дятлов (3—5 яиц; см. очерки по первому и второму виду там же). У D.major и других видов с доминированием самок (сирийский D.syriacus) последние участвуют в гнездостроении, потом и в выкармливании птенцов много меньше самцов.
Чем хорош этот вид как модельный объект?
Иными словами, одиночная территориальность «больших» пёстрых дятлов «живёт» с августа до начала—середины апреля. Чуть позже, к концу апреля они переходят с семян хвойных на насекомых, пьют берёзовый сок и т. д. (отдельные случаи раздалбливания шишек, в т.ч. упавших, фиксируются в мае и в июне7). При невозможности найти нужное дерево на объединённом участке (что чаще всего происходит именно в лучших зимовочных биотопах) пара распадается. Птицы исчезают с него и перемещаются в поисках подходящих гнездовых биотопов, иногда далеко, хотя до сих пор частота участия в весенней дисперсии в популяции, её дальность в разных обстоятельствах изучены плохо, и вовсе никак — выбор биотопа при оседании.
Впрочем, территориальные отношения заканчиваются в обоих случаях. Социальная жизнь, кипевшая в группировках осенью и зимой, прекращается полностью. Подавляющее большинство поселений (в т.ч. 3 из 5 бывших предметом долговременных наблюдений, все с наибольшими значениями плотности, и максимумом её межгодовых колебаний) в гнездовой период пустеют. Большая часть резидентов исчезает с территорий в конце апреля—начале мая и гнездится в иных местах и в иных биотопах.
Это значит, что каждый следующий «запуск» внегнездовой территориальности в конце лета независим от предыдущих. Персонализованные отношения, могущие сложиться между владельцами, «обнуляются» ежевесенне. В августе-сентябре они «строятся» заново, даже когда до трети занявших осенне-зимние территории взрослых — те же, что в прошлые годы (включая год назад сеголетками, отличимых до конца сентября по красному темени).
Статистический анализ выборки в 186 «повторных» занятия территорий в трёх плотных поселениях самцами и 172 — самками (за все годы наблюдений) показывает, что в следующий сезон (-ы, до 4-х) оно независимо от расположения территорий в прошлом. Даже более: когда часть из них отслеживали с момента прибытия8, место, где они держатся, «зародыш» участка, размещался случайно по отношению к территории (-ям) прошлых лет.
Ежегодный разрыв преемственности территориальных связей во внегнездовых группировках при сильной привязанности к ним резидентов9 делает больших пёстрых дятлов отличным модельным объектом в исследовании наиболее общих механизмов коммуникации, сперва «порождающих» разнокачественность особей (т. е. социальную стратификацию и сопряжённую с ней дифференциацию поведенческих ролей) в последовательных взаимодействиях между ними, потом преемственно развивающих эту структуру связей в течение осени и зимы… до гарантированного «обрыва» следующей весной.
Подобные механизмы пока что малоизучены, в т.ч. их универсальность их пока под вопросом, оживлённейшие дискуссии этологов на сей счёт частью изложены в монографии «От стимула к символу», ч.2 (где см. сравнительную таблицу концепций коммуникации). В любом случае они «делают» из однотипных отношений дифференцированные, из однородных особей разнокачественных: вопрос в том, как и «какими средствами»? Используя удобную здесь социобиологическую терминологию, можно сказать, что это проксимальные механизмы, противопоставленные ультимативным (объясняющим факторы эволюции, приведшие к такого рода эффектам). Но каковы бы ни были механизмы «создания» разнокачественности, они противопоставляются персонализованным (ролевым) отношениям между особями. Во втором случае птицы разнокачественны с самого начала, что «проявляется» и «учитывается» в социуме за счёт их знакомства друг с другом, выработавшего «привычку» к контактам на известных отрезках границ с известными соседями. Тогда специальных коммуникативных механизмов не нужно вообще, всё складывается само собой, достаточно птицам какое-то время жить по соседству… и чем дольше живут, тем сильнее эффект.
И именно у больших пёстрых дятлов такой способ становления разнокачественности исключён. По крайней мере, этих влияний нет и не может быть в первые 3—5 недель после занятия резидентами осенне-зимних участков (в августе, иногда и с конца июля). Этот период — самый значимый для социальной стратификации в новообразующемся поселении: задаёт её скорость на всё время зимовки и общий размах дифференциации поведенческих ролей у особей разного статуса. Как дальше увидим, в «точке 0» жизни поселения владельцы территорий в известном смысле однородны, независимо от пола и возраста10, в первую очередь по уровню агрессивности и её выражению в ритуализованной форме и напрямую, во вторую – по активности во взаимодействиях с соседями, готовности самому их инициировать или, наоборот, вступать во взаимодействия с соседями, не уклоняться.
Дальнейшая диверсификация особей по этому показателю и другим, из него вытекающим (успеху охраны собственного пространства, контролю перемещений и вторжений соседей, частоте собственных попыток захватов периферии чужого участка и пр., рис.11, 3, 5) «делается» историей побед и поражений в конфликтах друг с другом. В первые дни после образования группировки она более-менее одинакова у разных владельцев, как и их уровень агрессивности, оцениваемых по встречаемости прямых действий нападения и бегства (клевков, вытеснений, отскоков и пр.) в отслеживаемых потоках активности (рис.9). Потом – в следующие недели и месяцы – они всё больше разнятся между особями, причём у каждой из них сперва изменяется первое, потом второе (рис.13-14).
Ежегодный «перерыв на размножение» облигатен именно в плотных поселениях, >15 особей/км2. При более низкой плотности часть дятлов (иногда значительная), гнездится прямо на осенне-зимних участках или в радиусе до 500 м вокруг благодаря «вкраплениям» мелколиственных пород в хвойные древостои. Правда, и здесь социальные связи с соседями (предполагающие активное и направленное инициирование взаимодействий «с обеих сторон», о чём ниже) полностью рвутся с началом гнездостроения. Птицы лишь реагируют (агрессией) на появление других дятлов (своего, в ослабленной форме — и близких видов) на нарушение дистанции 15—20 м вокруг места пребывания. С началом насиживания она сокращается до ствола гнездового дерева, реже метра-двух вокруг дупла.
В плотных поселениях, наоборот, социальная стратификация и разнокачественность особей гарантированно возникают de novo каждую осень. Мы исследовали, как именно это происходит. Каждый осенне—зимний сезон образует как бы одну повторность в выборке «жизней» социумов шести пространственно разграниченных поселений суммарно за 38 прослеженных нами сезонов от августа по декабрь, каждый раз разворачивающихся «с нуля». См. данные, использованные для анализа,в табл.1:
| Данные | Общий объём |
| Годы наблюдений | 1984-1986, 1989-1998, 2002-2009 гг., 2011-2017 гг.. «Жизнь» плотных поселений прослежена от образования до образования пар весной на протяжении 47 сезонов, разреженных — 23. |
| Прослежена судьба | 516 особей в 6 модельных группировках (267 самцов, 249 самок, из них 84 самца и 108 самок здесь впервые отмечены сеголетками) |
| Обычный период пребывания особи в составе группировки | 2-6 осенне-зимних сезонов, не всегда подряд (по 2 раза — 7 сезонов, по 1 — 8, 9 и 10) |
| Процент гнездящихся на осенне-зимних территориях и не далее 500 м от них | 20-45% в разные годы |
| Радиус контроля за переселениями птиц между зимними группировками | «Лосиный остров» — 10-16 км; Павловская слобода – 5-7 км; пойма р.Нерской и Аминьевский лес, Грызловская дубрава, Пироговский лесопарк, ГБС — нет. Три последних — разреженные поселения, наблюдаемые относительно кратковременно |
| Объём данных, использованных для анализа | Территориальные конфликты – 16311; Дистантные угрозы – 23180; Маркировка территории – 29372; Патрулирование границ территории (периферии участка) – 9145; Ухаживания «внутри» территориальных конфликтов – 671; Ухаживания на этапе поиска партнёров — 6380; То же, на стадии консолидации пары — 5149. |
| Основные методы наблюдений | Картирование значимых событий социальной жизни особей; Хронометрирование всех действий особей в их последовательности |
| Из анализа исключены | контакты с не установленным исходом, с пробелами в регистрации >5 с., с потерей птицы из виду на >25% времени стычки |
Складывание территориальной структуры и анизотропность пространства плотного поселения
При занятии одиночных территорий в августе—сентябре дятлы сперва пробуют закрепиться в самых плотных поселениях данной местности, хотя там удержать их труднее всего — конкуренция ожесточённа, в выборке поселений доля «неудачников» среди прибывших туда птиц растёт быстрей роста плотности. Анализ показывает обратную зависимость последней от плотности: когда она <15 особей/км2, вселенцы, не сумевшие занять территорию, просто отсутствуют (другое дело, что в следующие месяц-два — да и всю зиму — она может серьёзно сократиться или быть вовсе потерянной, если дятел не защитил её центр от соседских вторжений). Возникает «естественная» граница плотных поселений, где владельцы активно поддерживают жёсткую территориальность всю осень и зиму11. Последняя и распределяет птиц между поселениями данной местности (рис.2). В августе-сентябре дятлы пробуют занять территории в возникающих плотных поселениях, а потом удержать охраняемое пространство вопреки постоянным попыткам соседей и/или новых вселенцев захватить его периферию. Там для них привлекательнее всего, но и трудней удержаться: «неудачники» исчезают оттуда и появляются в более дальних и разреженных поселениях, занимая территории там. Как минимум те 101, которых — в 1993-2007 гг. — мы обнаруживали там довольно случайно; 42 из них сменили 2 и более поселений, не сумев закрепиться там, и в следующие разы выбирали более разреженные.
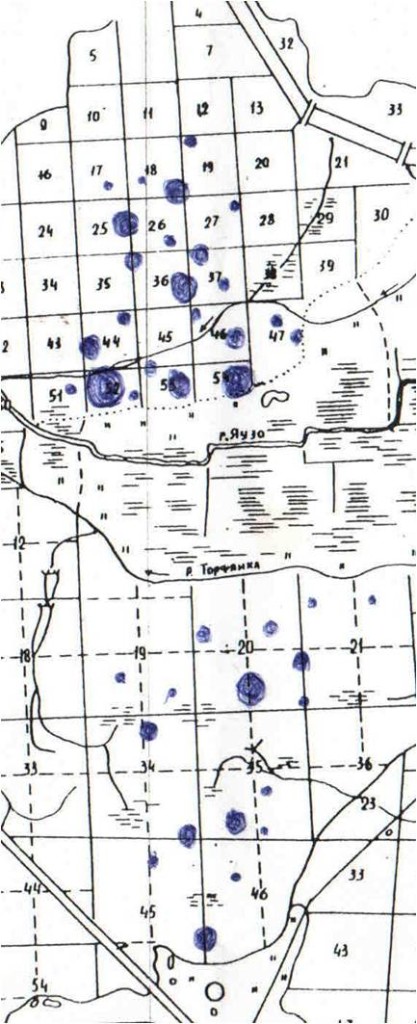
Рис.2. Пространственная картина поселений больших пёстрых дятлов на учётном маршруте в загородной части «Лосиного острова» по данным картирования в октябре—декабре 2003 г.
Примечание. Каждое пятно включает минимум две-три территории, маркируемые и охраняемые хотя бы в центре, резидентные одиночки и бродячие особи не отмечены, размер пропорционален плотности поселения.
Вторгаясь на чужие участки и обороняя собственные от вторжений соседей, владельцы всю зиму пробуют «переделить в свою пользу» общее пространство поселения. Они также живут «под давлением» активных вселенцев, пробующих закрепиться в местах, охраняемых слабее всего — на краю группировки или на стыках территорий. Если это удастся, «зародыш» участка (15-20 м вокруг основной «кузницы» захватчика) дятел расширяет до обычных размеров с одновременной интенсификацией охраны, рекламирования пространства и патрулирования границ.
Адаптивный смысл этих процессов до сих пор не изучен, и интересно выяснить, есть ли он вообще, не удовлетворяясь стандартными объяснениями вроде монополизации пространства и/или ресурсов. Тем более столь высокая плотность и жёсткая территориальность равно отсутствуют у близких форм — белокрылого D.leucopterus, сирийского D.syriacus и белоспинного дятлов D.leucotos. Первые два настолько близки, что образуют общий надвид с D.major; но и третий им близкородственен, по данным молекулярной систематики12. На «дне» падения численности в Финляндии он интенсивно гибридизировал с D.major, и гибриды были плодовиты. Т.ч. эта особенность скорее функциональна, чем исторична, поэтому адаптивное объяснение предпочтительней.
Неуспешность охраны территории — учащение поражений, снижающих вероятность победы в n последних стычках ниже некого минимума, за чем следует сокращение её площади ниже предельных 2000–2400 м2 — ведёт к её потере. В среднем по выборке плотных поселений «нижняя планка» успешности охраны пространства зависит от напряжённости территориальной конкуренции (частоты нарушений границ и попыток территориальных захватов), причём колоколообразно — сперва увеличиваясь с её ростом, а потом падая. Зависимость от плотности поселения много слабее и более косвенная. «Неудачники», не сумевшие закрепиться в поселении в первые дни после прибытия или удержать территорию в следующие дни, наполненные вторжениями соседей и вселенцев, составляют 60–95% птиц, привлечённых в плотные поселения в конце лета, и 15–45% – занявших участки и удерживавших их >3—5 дней, выстроивших там ночёвочное дупло. При прочих равных (общая численность вида, урожай семян в этом году и пр.) большая плотность поселения увеличивает процент «неудачников», «немедленных» и «отложенных», потерявших участки на протяжении осени и зимы. Второй ниже первого, но держится на одном уровне в течение всей осени и зимы, не снижаясь по мере «знакомства» птиц в группировке.
«Неудачники» дальше переместятся в ближайшие (~⅓ из них) и более дальние (~⅔) поселения, всегда менее плотные и с большими межгодовыми колебаниями численности. В поселениях, наиболее привлекательных для вселенцев, из года в год занятые участки сильно совпадают пространственно, даже когда их владельцы — разные особи. В поселениях, непривлекательных для вселенцев в конце лета-начале осени, но принимающих «неудачников» поздней осенью, наоборот, между территориями разных лет нет преемственности, почему их расположение в «сети» группировок и состав населения сильно варьируют по годам13. Территории там занимаются сильно позже, а их положение на местности (не только друг относительно друга) сильно меняется по годам.
Таким образом, поселения вида сильно дифференцированы по привлекательности для вселенцев и как следствие — по срокам занятия территорий, межгодовой преемственности состава, числа владельцев, расположения территорий между годами. Её степень зависит от интенсивности размножения вида в данном году, длительности позднелетней дисперсии и урожая семян; в общем случае дифференциация тем больше, чем выше численность вида в данной местности и интенсивней потоки вселенцев, не только перераспределяющихся внутри неё, но и приходящих извне, за счёт инвазий, обычных для более северных частей ареала14. Относительный «нуль отсчёта» в каждом из них, однако, один – первое занятие нескольких территорий в самых плотных поселениях прошлых лет (крупнейшие точки рис.2).
Самое интересное: резиденты предпочтительно расширяют занятое пространство всегда к центру поселения, не «равномерно во все стороны», и не «к периферии», хотя это самая неуспешная тактика: здесь сопротивление выше, а шансы на успех ниже (рис.3). Как и вселенцы, лишь претендующие на участок, они чередуют а) вторжения к соседям с последующей маркировкой/патрулированием новых границ15 криком «кик» с б) охраной «ядра16» собственной территории от их захватов. В сентябре и марте для маркировки птицы начинают использовать барабанную дробь, и учащают её использование после объединения участков партнёров в апреле.
I
II.
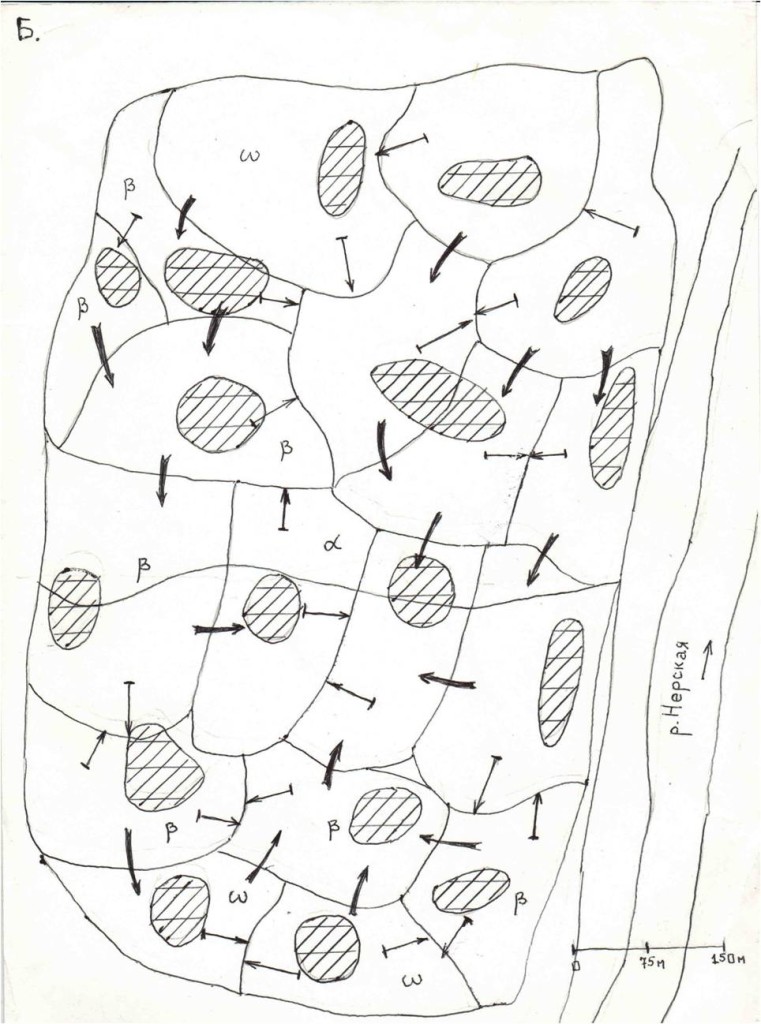
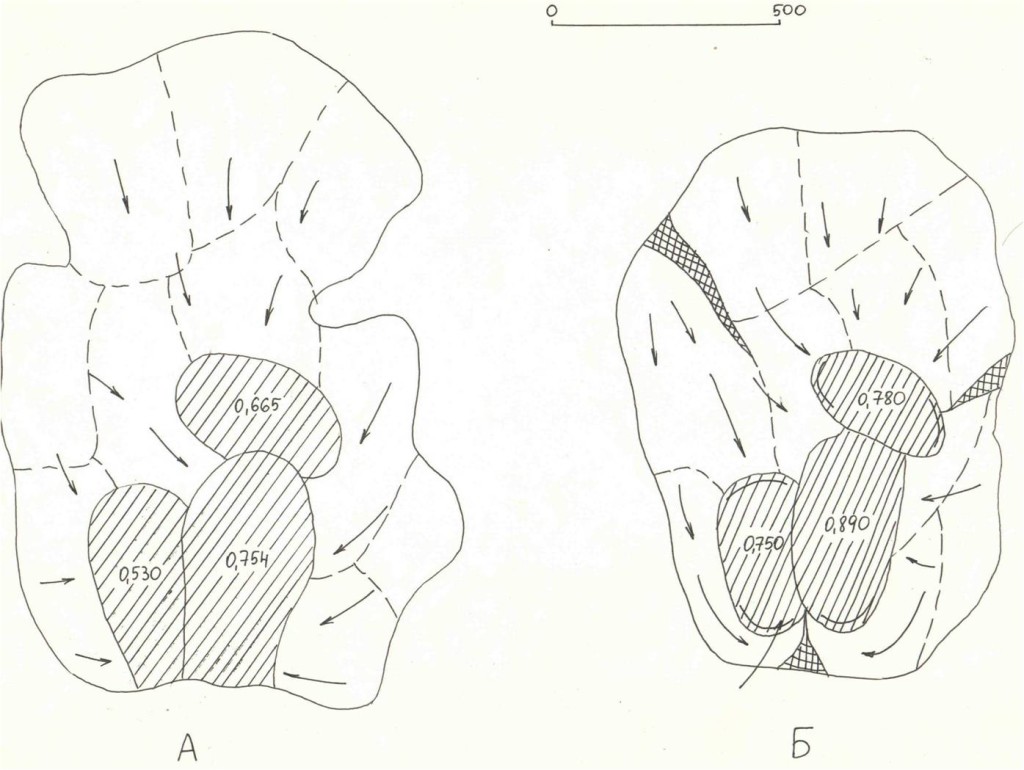
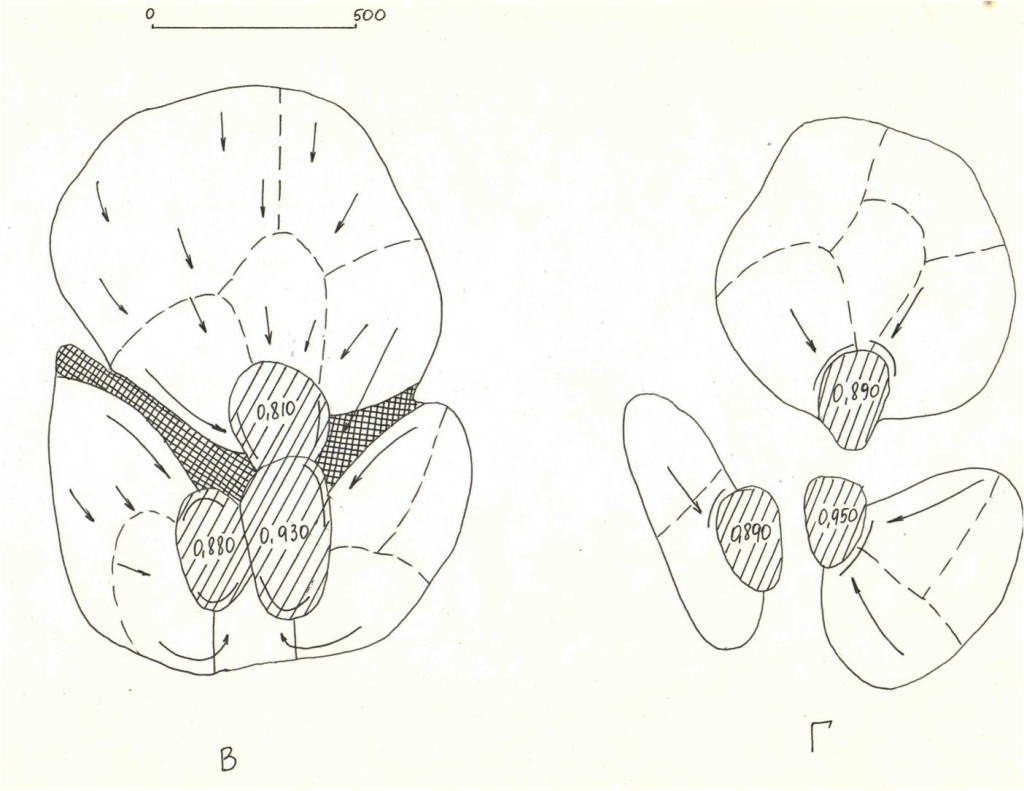
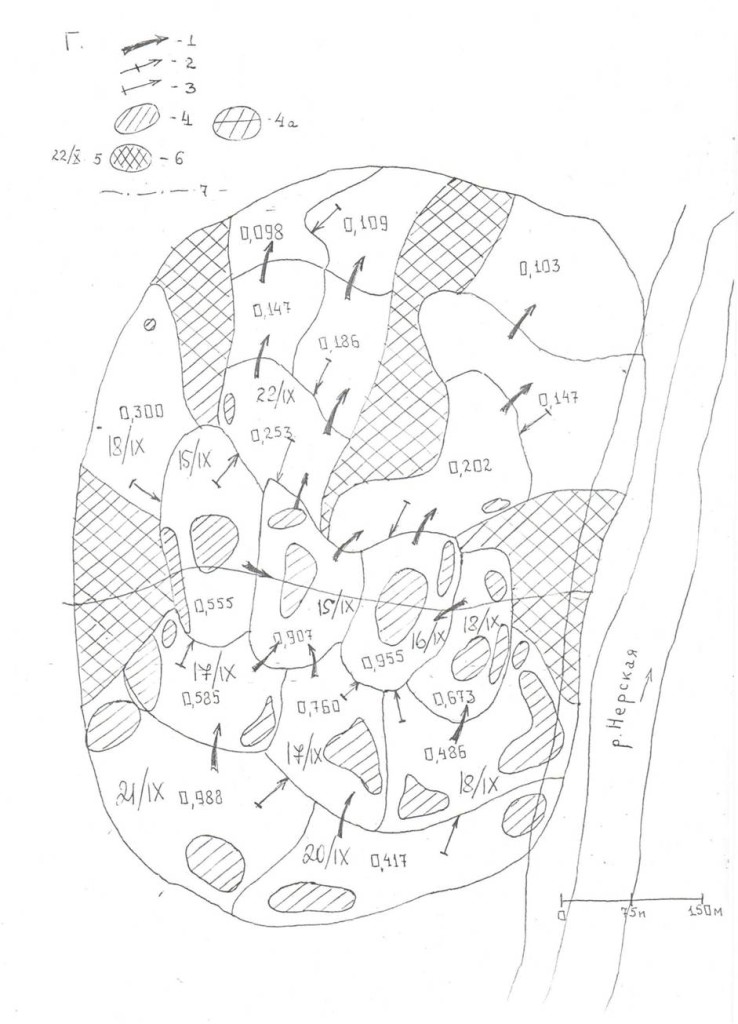
Рис.3. Последовательные изменений территориальной структуры плотных поселений в старовозрастном сосняке «Лосином острове» (кв.55, I) в августе—ноябре 1999 г. и перестойном ельнике у р.Нерской, с.Загряжское, в августе—ноябре 2002 г. (II).
Обозначения. I. Первые территории везде заняты к середине августа. А—Г — основные этапы трансформации территориальной структуры группировки к середине сентября, октября, ноября и декабря (сравни рис.11, а-ж). Стрелки — главнейшие направления территориальных захватов (>75% по времени и числу попыток), косая штриховка — пространство, охраняемое владельцами успешней всего (показан % побед в прошлых конфликтах), двойная штриховка — области, оставленные владельцами (переставшие маркироваться и охраняться, не говоря об эксплуатации, у D.major крайне локальной17). Жирные стрелки — доминантно-субординантные отношения между наиболее успешными резидентами.
II. 1 — попытки территориальных захватов, 2 — перемещения с маркировкой в «пустое» пространство, присоединяемое к участку, 3 — основная охрана периферии от попыток захватов, 4 — «ядра» одиночных территорий, 4а — области наиболее интенсивных конфликтов, α, β, ω соответствуют кластерам 1, 3, 2 рис.6. 5 — даты стабилизации границ, 6 —пространство, переставшее охраняться в связи с социальной стратификацией.
Чем большая доля конфликтов выигрывалась владельцем в прежние 3–5 дней сравнительно с соседями, тем чаще он демонстрирует «амбиции» — сам вторгается к ним в моменты, когда тем сопротивляться всего труднее: они сами проиграли конфликт с третьей особью. Или когда неудобней, т.к. вынуждает прервать долбление на взаимодействие. А дальше инстинкт «не пускает» вернуться к прежней шишке, «заставляя» лететь за новой и выбрасывать старую, где не использована значительная доля семян. В норме же дятел выбирает почти все семена18.
Для подобных вторжений и сопротивления им владельцы непрерывно отслеживают перемещения и взаимодействия соседей, чему помогает рекламирование территории криком «кик»: временной профиль волны «киканий» меняется в зависимости от того, победил дятел в недавнем конфликте или проиграл, и насколько «убедительно». И наоборот, «неудачные» относительно соседей дятлы преимущественно защищаются от попыток захватов, совершаемых в основном в те же «неудобные» для них моменты (Фридман, 1998 и рис.11). Это верно и для дистантной угрозы, когда захватчик так и не решается вторгнуться, обмен ритуализированными угрозами, часто в сопровождении выпадов, клевков и подскоков «в пустоту» идёт «через границу» участка на значительном расстоянии. Как и прямые конфликты, угрозы на расстоянии завершаются победой одной и поражением другой особи таким же образом и по тем же «правилам» (подробней см.рис.4, 8).
В выборке группировок, начиная с разреженных, времязатраты на это отслеживание растут с ростом плотности экспоненциально, как и на рекламирование «киканьем». Во время него резиденты концентрируют внимание на паттернах поведения, показывающих вероятный переход к территориальным захватам, вроде патрулирования границ — самого демонстративного поведения между конфликтами (т. е. без учёта собственно демонстраций «внутри» них). Оно часто осуществляется самой «яркой» и «красочной» из ритуализованных форм поведения вида — трепещущим полётом, т. н. «Flatterschwebflug» (Blume, 1968).
Или, наоборот, действиями противоположного облика: после рекламирования приграничного пространства птицы, держась максимально скрытно, проникают к соседям как можно «глубже» (считая по радиусу от границы до ближайшего «ядра» территории, где расположены «кузницы» владельца). Чем позже их застигнет владелец, тем большую площадь периферии они смогут «оттяпать», будет выиграют начавшийся при этом конфликт или группу конфликтов, следующих «впритык» друг за другом. Однако и риск проиграть растёт пропорционально «углублению», чем дальше от границы, тем ниже побуждение к нападению и готовность захватчика двигаться дальше.
Владелец, в свою очередь, следит за соседями и пресекает попытки захватов в зависимости от своей мотивации, с одной стороны, и её изменений в зависимости от дистанции до границы и/или центра участка. Дятлы, успешно защищающие пространство дятлы внутри него не показывают побуждения к бегству, почему много времени следят за соседями и интенсивно патрулируют полосу в 5-10 м у границы, а то и предпринимают рейды по чужой территории. Захваченных при этом соседей и/или вселенцев данные особи атакуют немедленно, не отступают поближе к центру участка, даже будучи атакованными обнаруженными захватчиками в ответ. Чем ниже доля побед владельца в территориальных конфликтах прошлого, тем «круче» падает готовность атаковать захватчика в направлении от «ядра» территории к периферии и, наоборот, быстрее растёт побуждение при встрече с ним отступать к центру.
Обнаружив нарушителей в периферийной части участков, подобные особи если и встают на пути их продвижения к центру, то быстро отступают под натиском (периодически, правда, пробуя контратаковать, «выталкивая» противника кнаружи участка). Возникают своего рода «качели» — дятлы мечутся взад-вперёд по градиенту «центр-периферия» территории, то теснят друг друга грубой физической силой, без сопровождающих демонстраций, то пробуют «обойти» оппонента или перелететь — или пройти, в зависимости от степени «плотности» контроля перемещений друг друга — в направлении прежних движений. Так продолжается от 1-2 мин до четверти часа, но рано или поздно птицы останавливаются в «точке равновесия», где продвижение захватчика к центру нейтрализуется атаками владельца в попытках его оттеснить, и переходят к организованному обмену демонстрациями и сопровождающими их прямыми действиями; это момент, когда социальный контакт перерастает во взаимодействие, исходы которого причинно связаны с выбором и употреблением демонстраций участниками и по результатам которого их поведение
а) дифференцируется количественно и качественно, в сторону взаимной комплементарности у разных особей, т.ч. они оказываются более связанными между собой, чем до этого;
б) направленно меняется в будущих взаимодействиях, в т.ч. с «третьими» особями, а не только друг с другом, в зависимости от исходов данного, т. е. они обладают последействием.
Место остановки «качелей» логично считать моментом равновесия антагонистических побуждений нападения и бегства внутри каждой из особей и одновременно между владельцем и захватчиком, что позволяет объективно оценить уровень агрессивности каждого и проконтролировать его измерения, основанные на действиях прямой агрессии, появившихся во время преследований и позднее, во взаимодействии. Из них важнее всего акты нападения разной амплитуды, от клевков до выпадов, подскоков, разворотов к противнику, и противоположные им акты бегства. Их интенсивность и соотношение между собой даст объективную оценку уровня агрессивности во взаимодействиях, неритуализованной (по действиям, появившимся в потоке активности между демонстрациями), так и ритуализованной (по исполняемым сопряжённо с предъявлением разных демонстраций).
В силу центростремительности территориальных захватов и центробежности обороны от них, дятлы вторгаются на чужие участки в основном через те сегменты периферии, что обращены к центру поселения. И наоборот, защищаются от попыток захватов преимущественно границы, «обращённые» к краям группировки. Благодаря этому уже через 1–2 месяца охраны участков (конец сентября–начало ноября), в центре поселения (функциональном, не обязательно геометрическом) автоматически оказываются особи с максимальной успешностью охраны пространства. Прочие не удержатся под «давлением» вторжений на «обращённые к центру поселения» части их собственных территорий (рис.3А, Б).
Так пространство поселения делается анизотропным. Его центр привлекательней всего для захватчиков и резидентов, включая самых периферийных и неуспешных: т.е. «качество» защищаемого и оспариваемого пространства падает центробежно, что управляет поведением тех и других птиц.
Охрана территорий и результаты конфликтов: положительный, отрицательный, нулевой
Пространство территориальной активности дятлов делится на охраняемое (в плотных поселениях — в границах участка), рекламируемое — в пределах функциональных зон, мест пребывания владельца и вторжений захватчиков, и регулярно обследуемое — в их поиске. Оно: 1) много больше используемого для кормления и 2) защищается от соседей и «активных вселенцев», независимо от пола и возраста, но не проходящих особей. Последние свободно кормятся и перемещаются по участкам владельцев, пока не пробуют стать резидентами. Тогда они останавливаются и, держась крайне скрытно, интенсивно отслеживают территориальную активность резидентов. Обычно они это делают вечером во время вечернего «всплеска» маркировки (1,5-2 ч до захода в ночёвочные дупла). На следующее утро претендент максимально интенсивно маркирует своё место пребывания (>60-90 «киков»/мин). При слабой охране, что обычно на «стыках» нескольких территорий, он также пробует захватить часть периферии у одного из владельцев, слабее откликнувшегося на его маркировку, в ответ на неё не появившегося для патрулирования и пр.
Отмечены три формы защиты участка от таких «покушений»
1а) просто силовым вытеснением. Иногда владельцу достаточно лишь появиться и тот ретируется, но бывают и раунды клевков с замещающими прыжками (англ. supplanting attack), при низком возбуждении лишь замещающие прыжки или направленное движение «пешком» по веткам в сторону нарушителя, и тот в конце концов убирается,
1б) если он упорствует, не остаётся на месте и даже прорывается вглубь территории19, птицы останавливаются и переходят к обмену демонстрациями, также сопровождаемому открытой агрессией, разного рода перемещениями и т. д. неритуализованными элементами поведения. Они появляются либо вместе с демонстрированием, и более-менее сопряжены с предъявлением каждой из основных демонстраций во времени, действия нападения или бегства вместе с соответствующей локомоцией образуют своего рода «выплеск», реализуемый в отношении противника дятлом, удерживающим демонстрацию, и продолжающийся ещё некоторое время после неё.
Также они появляются в паузах между демонстрированием, «заполненных» повседневной активностью с локомоцией, реже смещённым долблением или «сном» (а реже всего — смещённым «поеданием» снега). Кроме них, на противника воздействуют «киканьем» барабанными дробями (весной, реже поздним летом и осенью) и,
1в) дистантной угрозой — самой ритуализованной формой защиты. Здесь агонистические демонстрации (избираемые также и в той же последовательности, что в конфликтах) адресуются оппоненту, когда он у себя на участке, в десятках метров от птицы. Ею останавливают потенциальных нарушителей, когда они движутся к границе или по ней. Самый частый предиктор угроз — характер маркировки «пограничной полосы», см. далее.
К территориальному поведению также относятся
2) рекламирование места нахождения особи или более протяжённых районов внутри участка криком «кик», в периоды брачного возбуждения (конец сентября—начало октября, с конца января и далее) также и барабанной дробью. Верещанье маркирует рост возбуждения, вкупе со слабой агрессией, у владельцев при отслеживании перемещений соседей, когда их стрессирует их движение к границе и возможность нарушить.
3) Патрулирование границ обычным и/или трепещущим полётом. Наиболее возбуждённые дятлы также верещат перед или во время этого. Данный крик часто фиксируется перед встречей с потенциальным партнёром, когда он ещё воспринимается как нарушитель и возможный захватчик. Видимо, два сонаграфически различающихся варианта крика жёстко связаны с двумя конфликтами побуждений «с участием агрессии» — нападение-бегство и нападение-бегство-половое поведение.
Формы поведения 1)—3) составляют территориальную активность. Она сильно варьирует в зависимости от плотности: в зависимости от неё меняются выраженность 1)-3) (общее разнообразие, доля ритуализованных и специфических элементов — восьми территориальных демонстраций рис.4) и доля бюджета времени, приходящегося на эту активность. С ростом плотности частота, средняя длительность и интенсивность конфликтов также растут, и быстрее чем плотность, по экспоненциальной кривой. В сравнении с однотипной зависимостью для рекламирования она «запаздывает», но столь же «крута». В тех же координатах растёт доля ритуализированных демонстраций в конфликтах, дистантных угрозах и патрулированиях участков.
Таким образом, у больших пёстрых дятлов, как и других видов птиц, агонистические взаимодействия, в т.ч. территориальные конфликты, разрешаются именно обменом ритуализованными элементами, организованным по видоспецифическим или общим правилам, т.е. коммуникацией с демонстрациями-посредниками. И наоборот: попытки их разрешения прямой агрессией безрезультатны у всех видов, специально исследованных в данном аспекте20. Они ведут лишь к обмену клевками, выпадами и т.д. реакциями открытой агрессии до изнеможения или ранений участников, сильно затягивающемуся во времени и неизменно безрезультатному.
А.
Б.
Рис.4. Территориальные демонстрации большого пёстрого дятла, «расставленные» на градиенте территориальной агрессии (соответствует радиальному срезу участка от основной «кузницы» к границе и далее вглубь чужой территории) по наибольшей вероятности появления первой в конфликте, после того как владелец остановит захватчика, пробовавшего углубиться возможно дальше, и оба перейдут к обмену демонстрациями. №№1—4 – позы нападения, 5—6 — амбивалентная поза (угрозы), №№7—8 — позы подчинения. Прочие обозначения см. рис.4Б.
Обозначения. А. Внешний облик. Пунктир — «ядро» территории. Стрелки — элементарные движения, составляющие демонстрацию. Тогда они исполняются лишь все вместе, а исполнение жёстко сопряжено во времени, что формирует типический образ каждой демонстрации. См. подробнее Фридман В.С. От стимула к символу. Ч.2, рис.2. М.: URSS, 2013.
Б. Эффекты — частоты «автоматической» выдачи реципиентом движений атаки (1), бегства (2), остановки и перехода к демонстрированию (3) в ответ на демонстрации №№1№№1—8, иные демонстрации (Б), также встречающиеся при охране территории (в основном брачные в январе—марте), прочие элементы поведения, включая незавершённые и неточно исполненные демонстрации основном локомоции и экспрессивные реакции (В). n (предъявлений демонстраций №№1-8=354; 296; 307; 411; 438; 385; 300; 183. «Иные К2» — демонстрации иных контекстов, прежде всего брачные (n=1150 предъявлений), К1 – экспрессивные реакции, яркие, демонстративные, но не обладающие устойчивой формой (n=194), НК – демонстрации, исполненные неполностью или неточно (n=217), НР – прямые действия агрессии, бегства и пр. (n=288). Соотношение (1)—(3) характеризует каждую демонстрацию и закономерно меняется в ряду от №1 к №8, т. е. «вдоль» градиента территориальной агрессии (или «вниз» по нему).
Иными словами, августовское привлечение вторичным сигналом («киканье», другие звуки и образы, связанные с конфликтами, или следующие из их результатов) птиц, пытающихся занять территории, заметно опережает привлечение первичным сигналом (семена хвойных). Чем выше риск лавинообразного роста затрат на конфликты в гипотетическом случае столкновений при свободном перемещении в пространстве вокруг выбранных «баз операций» («кузниц» или ночёвочных дупел), тем это опережение больше, а «разгораживание» ранее общего пространства на индивидуальные территории во вновь возникающих поселениях происходит быстрее, как и дальнейшие этапы социальной стратификации (рис.11). Так птицы переводят времязатратную и опасную силовую борьбу в плоскость организованного обмена демонстрациями, т.е. коммуникации, результаты которой определяют выигравшего и проигравшего в конфликте не только для них двоих, но и поселения в целом.
Вообще, вторая особенность больших пёстрых дятлов — исключительный полиморфизм социальной организации во внегнездовое время (близкие виды, напротив, однообразны). Развитость территориальности «пробегает» все значения от 0 через промежуточные состояния (1—3) до максимума (4). «Нуль» здесь — неохраняемые участки обитания, одиночные, редко парные, соседи свободно посещают кузницы друг друга, но без агрессии, в исследовательских целях. 1 — поддержание индивидуальной дистанции: охрана более или менее протяжённого пространства сначала вокруг самой особи, потом (при увеличении плотности) вокруг её основной кузницы (2). 3 — охрана более или менее протяжённого пространства в центре участка («ядро»). Его периферия не охраняется и используется соседями совместно (режим 0). 4 — жёсткая охрана всего используемого пространства, с чёткими линиями границ. Одно приближение к ним соседей, настораживает владельца, если перерастёт в нарушение, то ведёт к разным формам угроз и агрессии. Их эскалация продолжается, пока не разрешится конфликтом в месте, детерминированном отношением уровней территориальной агрессии резидента с захватчиком. 5 — вышеописанное сочетается со своеобразным доминированием более успешно охраняющих территорию дятлов над неуспешными21. Первые («доминанты») имеют доступ на участки вторых («подчинённых») без попыток остановки, атаки и пр. Их присутствие не ведёт к «неприятным последствиям» в виде территориальных захватов именно потому, что при встречах с высокостатусным «нарушителем» «подчинённые» дятлы не «запускают» территориальное поведение.
В противоположность нарушениям со стороны соседей сравнимого статуса, здесь они ведут себя скромно и осторожно, точь-в-точь как вселенцы в первые дни после прибытия: при контакте с «доминантом» немедля распластываются с выдвинутыми рулевыми, т. е. сразу же демонстрируют позу №8, которой проигравшие особи завершают обмен демонстрациями в территориальном конфликте (там она маркирует остановку сопротивления, см. рис.7 и описание ниже). Но только в режиме 5: в режиме 4 и прочих даже самые неудачливые владельцы неизменно останавливают нарушителей и вступают в обмен демонстрациями. Проигрыш последнего и ведёт к неуспешности охраны пространства, отнюдь не меньшая агрессивность или физическая сила сравнительно с оппонентом (см. однотипный пример с изгнанием сеголеток с гнездового участка).
Все формы территориальности полностью развиты в плотных поселениях (>15 особей/км2). Там границы проходят с точностью до метра (или одной стороны ствола), дистантные угрозы и патрулирование границ запускаются даже в ответ на его патрулирование или при длительной маркировке соседа вблизи них. Поэтому на чужой территории захватчик ведёт себя крайне осторожно, чтобы, оставшись незамеченным, как можно ближе продвинуться к центру участка. Напротив, владелец, если успешен в конфликтах и/или только что выиграл прошлые, перехватывает нарушителей как можно ближе к границе. Если же неуспешен и/или недавно их проиграл, то отступает поближе к центру и уже там останавливает нарушителя.
В обоих случаях после серии «маятниковых» перемещений по радиальному «срезу» участка (размах их и способ перемещения – прыжками в кроне или перелётами – задан уровнем общего возбуждения обоих), дятлы останавливаются и вступают в организованный обмен демонстрациями, т.е. собственно в территориальный конфликт. Он может завершиться успешно: устойчивым разделением особей на выигравшего и проигравшего, и «проекцией» его на «спорную» территорию — её или удалось отстоять, или она переходит к захватчику. Другой вариант — он безрезультатен: в какой-то момент разлетаются с потерей интереса друг к другу, или взаимодействие «срывается» в драку, долгую и опасную для обоих, без возможности дифференцировать роли. Динамика внутренней структуры территорий по мере «накопления» индивидуальных историй побед и поражений у особей двух противоположных категорий — обычно выигрывающих vs проигрывающих конфликты показана на рис.5.
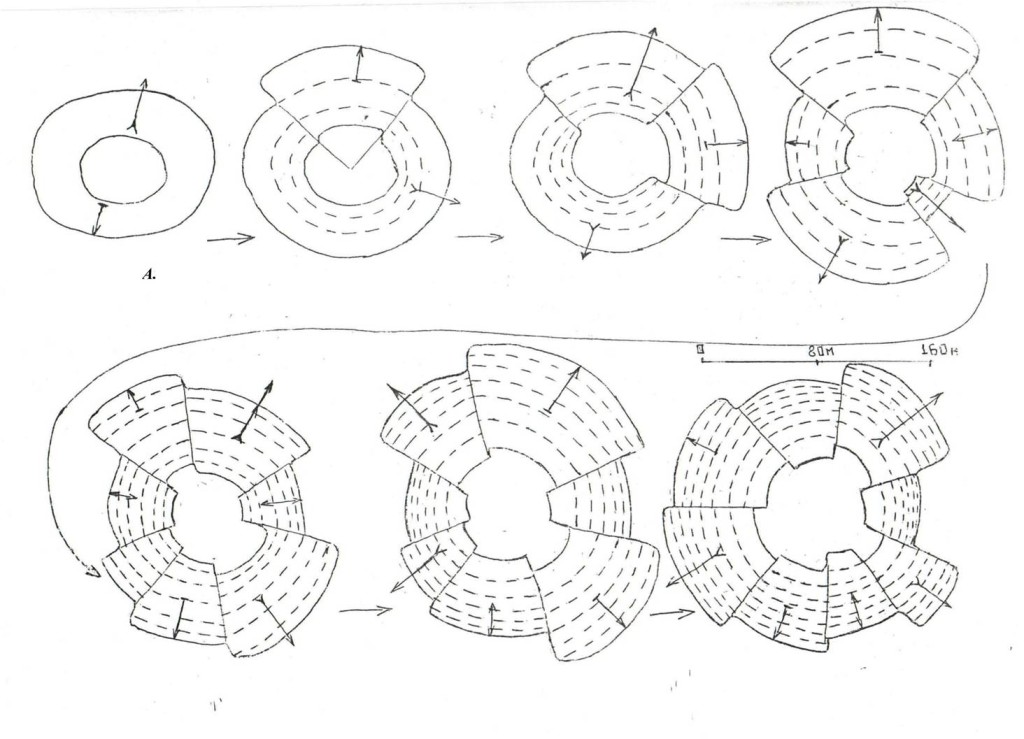
Рис.5. Динамика формы и структуры территорий по мере выигрывания и проигрывания конфликтов. Схема. А. «Меньшинство» особей кластера 1 (больше вторгаются и контролируют чужие конфликты).
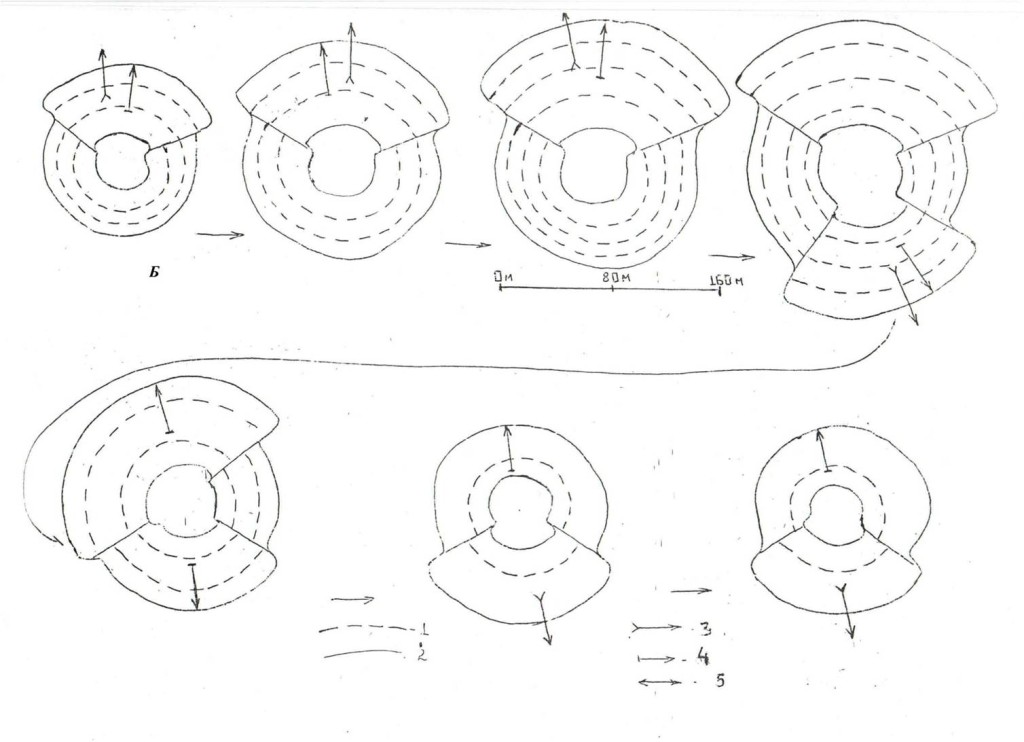
Рис.5. Динамика формы и структуры территорий по мере выигрывания и проигрывания конфликтов. Схема.Б. «Большинство» особей кластера 2 (лишь защищаются от вторжений). Каждый показанный сектор – область конфликтов с одним из соседей, сокращающийся с проигрышами и растущий кнаружи с выигрышами конфликтов, линия внутри него – «ядро» территории.
Обозначения. 1 – основные дистанции, на которых идёт маркировка после конфликтов, 2 – границы территории и «ядра», 3 – основные направления вторжений к соседям, 4 – основные направления защиты от их вторжений, 5 – места особенно продолжительных пограничных конфликтов (до 1,5–2 ч.): когда птицы активно перемещаются туда и обратно между участками, а изгоняемый и защитник своей территории многажды меняются местами.
Примечания. Схема показывает типические изменения территорий у птиц кластера 1 (особенно самец 5) и кластера 2 (особенно самка 15) осенью 1994 г. в поселении кв.55 «Лосиного острова».
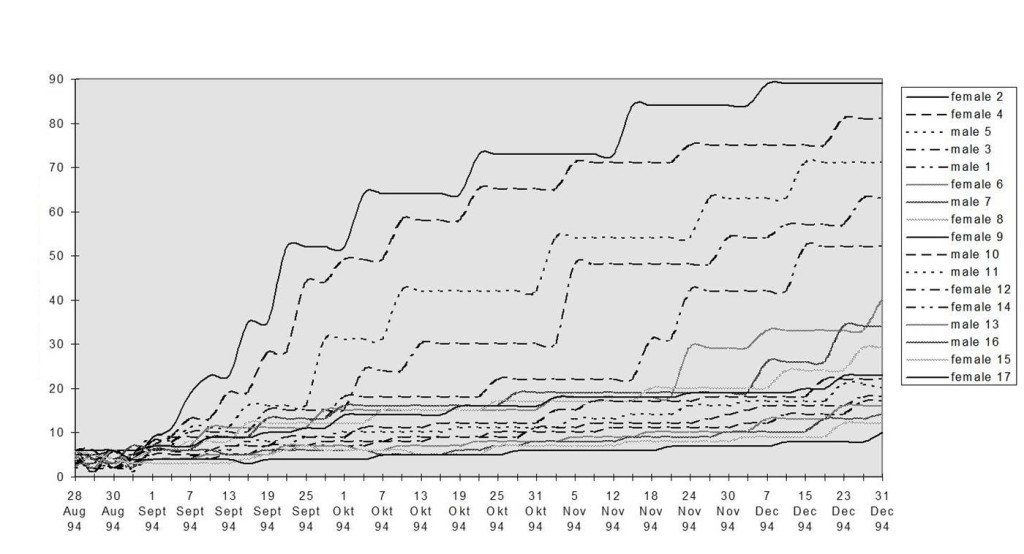
Рис.6. Социальная дифференциация в плотном поселении «Лосиного острова» осенью—зимой 1994 г. Обозначения. Ось Х — даты, У — успешность охраны пространства, % побед в 100 последних конфликтах (неделя непрерывной охраны). Кластеры 1 и 2, см.рис.5, 3 — см. ниже.
Организация обмена демонстрациями во взаимодействиях, прежде всего временная («синтаксис»), инвариантна в широких пределах изменений обстоятельств контекста, в которых они случаются между дятлами (рис.12). Она не зависит ни от частоты социальных контактов (растущей в степенной зависимости от плотности), ни от вероятности их перерастания во взаимодействие — территориальный конфликт или угрозу, и начала коммуникации (меняющейся примерно также, но также обратно зависящей от времени с момента прошлого нарушения), ни от интенсивности реакций особей во время собственно взаимодействия (измеренной скоростью смены отдельных поведенческих единиц, долей ритуализованных форм поведения и пр.). Эта организация подчиняет себе выбор и использование демонстраций участниками, определяя на всём протяжении процесса, какие демонстрации предъявляются, в какой последовательности, на какие сменяются22. «Синтаксис» взаимодействий территориальной агрессии больших пёстрых дятлов один и тот же независимо от того, неподвижны участники «обмена», или преследуют друг друга на стволе и ветвях «в пешем строю», либо гоняются друг за другом, используя полёты-прыжки. Интенсивность их локомоции участников везде пропорциональна уровню общего возбуждения и не влияет на его ход и/или результаты. То же фиксируется в агонистических взаимодействиях тех видов птиц, что специально исследовались в этом отношении. См. данные по бирюзовой акаре Aequidens rivulatus ((Maan, Groothuis, 1997; Maan et al., 2001), другой цихлиде Nannacara anomala (Hurd, 1997; Brick, 1998), «вращениям» лебедей-шипунов Cygnus olor (Lind, 1984), «дуэлям кивания» тетеревов Lyrurus tetrix (Лысенко, 1985, 1987) конфликтах домашних кошек Felis catus (Мовчан, Чернышёва, 1988), бойцовых рыб (Simpson, 1968), серебряных белоглазок Zosterops lateralis (Kikkawa, Wilson, 1983, 2002; Kikkawa, 1987) и т.д. видов. Их обзор и анализ см. «От стимула к символу», Ч.1—2.
В поселениях средней плотности (10—15 особей/км2) охрана периферии много менее тщательна (режим территориальности 3). Нарушителей «перехватывают» лишь в «ядре», на периферии их атакуют, лишь когда (и если) те начнут маркировку. Возможно совместное пребывание там, отдых и/или кормление соседних владельцев c поочерёдным использованием одних и тех же «кузниц».
В разреженных поселениях дистантные и «превентивные» (патрулирования) формы охраны вообще не встречаются (режимы 0—2). Охраняются лишь ближайшие окрестности «кузниц», используемых птицей здесь и сейчас. Нарушители беспрепятственно движутся в центр участка, владелец редко окрикивает их и атакует только в «конце пути», при посадке прямо на «кузницу» резидента или попытке маркировать территорию рядом с ней. Охрана пространства здесь так неустойчива, что 2/5 нарушений кончаются «ничем» — сосед останавливается и начинает кормиться, будто «забыв» о территориальных претензиях. В этом случае его и не трогают, через какое-то время он спокойно улетает к себе.
Вернёмся к плотным поселениям, где территориальные отношения всего сложнее, разнообразней и интересней. Последующие конфликты владельцев участков с соседями и вселенцами «отталкиваются» от сложившейся структуры как праймера, изменяют и переделывают её сообразно накапливающейся индивидуальной истории конфликтов, выигранных и проигранных, начавшихя ближе или дальше к границе, — и часто до неузнаваемости (рис.5).
Вселенцы, прибывшие в поселение, т. е. «несведующие» в его территориальной структуре, после прибытия не перемещаются как хотят, но «выявляют» эту последнюю. Первый день они держатся незаметно, всячески уклоняются от встреч с резидентами, но не обращают внимания на «проходящих» особей (контакты с последними неизменно мирные, в августе-сентябре 2006 г. из 84 подобных случаев с 19 птицами отмечено только 2 стычки, в неожиданных встречах «нос к носу»).
Весь день, и особенно вечер, они держатся крайне настороженно, стараясь быть незаметными, двигаются в районы наиболее интенсивной маркировки или конфликтов, активно отслеживают то и другое. После чего останавливаются в слабее всего охраняемых частях поселения (23 раз наблюдали ночёвки новоприбывших, 13 из них — на ветвях деревьев, 6 — начали выдалбливать ночёвочное дупло, на что тратят 2-4 дня, 4 раза — ночевали в дуплянке).
На следующий день они сразу же, безо всяких «проб и ошибок», вторгаются на периферию участков тех особей, кто по сравнению с прочими охраняет пространство наименее успешно, «вообще» или часто проигрывал конфликты вчера, примерно в равной пропорции. В паузах между конфликтами с их владельцем вселенцы кормятся и активно рекламируют своё местопребывание, обычно в 25-40 м от них; там же возможны конфликты с резидентами, пробующими их изгнать. Это один из примеров т. н. «подглядывания» (англ. eavesdropping behaviour23), присущего всем резидентам и активным вселенцам, но не отмеченного у «проходящих» особей: те вполне безучастны и к рекламированию, и к территориальным конфликтам других дятлов. Напротив, первые две категории птиц, когда сами не участвуют во взаимодействиях, то активно отслеживают течение чужих, их исходы и/или демонстрации, употреблённые при их достижении24. По результатам слежения они корректируют поведение в будущих взаимодействиях в сторону большей успешности демонстрирования (что зависит от последовательности выбора демонстраций, моментов их предъявления и смены — см. далее), выбирают «наилучшее время и место для них», и «наиболее удобного» противника. Поэтому чаще «подглядывающие» особи выигрывают территориальные конфликты и дистантные угрозы тем чаще, чем больше «подглядывают», а интенсивность последнего пропорциональна уровню территориальной активности данной особи, прежде всего маркировок и патрулирований границ, и частоте конфликтов её соседей.
Действительно, «подглядывание» облигатно для резидентов, уже включённых в коммуникативную сеть группировки, и недавно прибывших птиц, занимающих там территории — но неизменно отсутствует у «проходящих», только кормящихся там без каких-либо территориальных амбиций. Понятно, это свидетельствует о сигнальной роли во взаимодействиях отдельных единиц-демонстраций или, мягче, включающего их поведения, и возможность использования сигнальной информации первых или вторых для «опережающего отражения» происходящего с выгодой для наблюдателя, непосредственно в нём не участвующего.
Неслучайно подобные данные не упоминаются этологами, считающими, что специфический результат коммуникации создаётся неспецифическими средствами, а ритуализованные формы поведения участников лишь неспецифически возбуждают их25. Нами показано, что у D.major демонстрации видового репертуара выступают предикторами потенциально возможных исходов взаимодействия, «по которым» участники корректируют собственные модели поведения, состязаясь друг с другом в этой корректировке. Конфликты неизменно выигрывают особи, корректирующие их лучше противников, но никогда — просто более возбуждённые, или сильнее воздействующие на оппонента теми же демонстрациями как релизерами, см. ниже.
Однако демонстрации таковы лишь «в своём» контексте общения, не в иных, где они регулярно встречаются из-за «смешанности» мотиваций животного, образующихся конфликтом альтернативных побуждений. Для 8 поз рис.4 это контекст охраны территории, но не сближение потенциальных партнёров в феврале—марте, где возможно появление данных поз из-за восприятия птицами друг друга как захватчиков (рис.7). Для 9 демонстраций рис.1 всё наоборот — они выступают предикторами будущего течения взаимодействий и поведения участников только в ухаживании, но не в других контекстах, хотя «вкраплены» во взаимодействия охраны территории, много реже — дистантной агрессии, если те следуют сразу после ухаживаний или между ними.
А.
Б
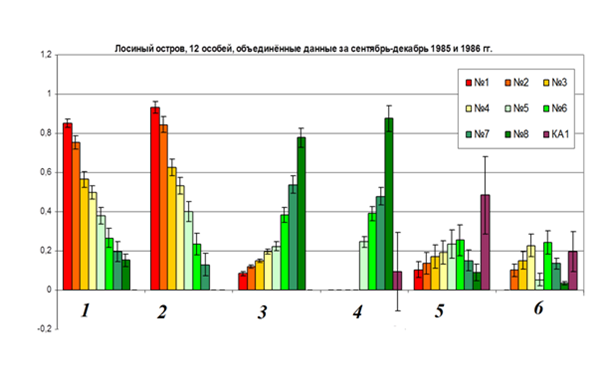
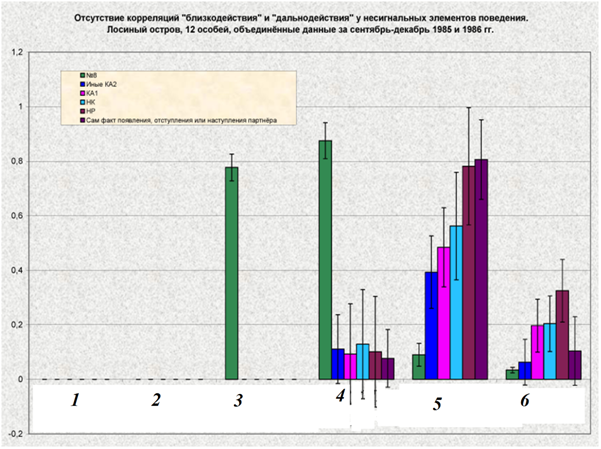
Рис.7. Корреляция «близкодействия» и «дальнодействия» 8 демонстраций территориальной агрессии (А) и других форм поведения, отмеченных во взаимодействиях (Б).
Обозначения. Ось У – вероятности исходов немедленных двигательных ответов на употребление разных форм поведения и наступления соответствующих им исходов взаимодействия в целом. 1 — вероятность победы демонстратора во взаимодействии, обусловленной данным сигналом, см. ниже; 2 – вероятность отступления/бегства после предъявления сигнала, 3 — вероятность поражения во взаимодействии, 4 — вероятность отступления или атаки после предъявления сигнала, 5 — вероятность «сбоев» процесса обмена демонстрациями (с наступлением паузы, заполненной локомоцией, смещёнными реакциями и пр.), 6 — вероятность срыва взаимодействия в целом (нулевой или отрицательный исход, см.ниже). КА1 – экспрессивные реакции.
Примечание. «Близкодействие» демонстраций — непроизвольный двигательный ответ реципиента, реализующийся после их предъявления независимо от других действий реципиента, демонстративных или нет, одновременно с ними и на их фоне, с определённой вероятностью. Последнюю определяли по частоте разнотипных ответов, если гипотезу о закономерной связи выдачи каждого из них с предъявлением данной демонстрации подтверждена дисперсионным анализом. Для 8 демонстраций территориальной агрессии таковы 3 вида ответов: атака, бегство и остановка с переходом к демонстрированию (F = 5,88; df1=7; df2=4523 предъявлений; P<0,01, хронометражи взаимодействий n=17 птиц из «Лосиного острова» за октябрь 1996-январь 1997 г. поселений), для 9 ухаживательных — продолжение токования в следующий период времени, сравнимый по длительности с предыдущим, включая более тесное сближение птиц vs спаривание vs атака vs уход от попыток сближения (F = 4,9; df1=8; df2=6744 предъявлений, P<0,01; хронометражи взаимодействий 33 птиц из «Павловской слободы» за февраль-апрель 1991, 1992 и 1993 г.). Демонстрации как фиксированные единицы поведения, обладающие типологически определённой формой, устойчиво дифференцированной от «фона» прочей активности и от форм других демонстраций, представляют собой сериальные элементы инстинкта (территориальной агрессии), кооперативно реализующегося в конфликтах резидентов с захватчиками, в той мере, в какой их следование во взаимодействии закономерно и предсказуемо не только для наблюдателя но, главное, для «подглядывающих» соседей. Т.е. в очень высокой: происходящее хорошо соответствует диалоговой модели коммуникации классических этологов (о ней см. «Взлёт и падение сравнительной этологии»; «От стимула к символу», рис.4, см.34, 71, 542 и т.д.). Каждая из употреблённых дятлами демонстраций — специфический актор данного «диалога», почему точно соответствует этологическому понятию социальных релизеров, предъявляемых «должным» образом в «должном» месте взаимодействия и «движущих» это последнее своим специфическим эффектом на следующий этап.
Как выглядит территориальное поведение больших пёстрых дятлов?
Успешными можно назвать взаимодействия, где участники, исходно равные между собой, в ходе взаимодействия устойчиво дифференцировались по поведению не только «здесь и сейчас», но и достаточно отдалённое будущее, включая поведение в будущих территориальных конфликтах. Первое объективно определяет выигравшую и проигравшую особь, не только «для нас», но, главное, для «зрителей» взаимодействия, «подглядывавших» за его течением и по завершению дифференцированно реагирующих на обоих птиц. Второе показывает последействие победы первого и поражения второго дятлов в территориальном конфликте, тем более долговременное и устойчивое, чем выше эффективность обусловивших их демонстраций (рис.12). Т.е. имеют место быть дифференциация поведенческих ролей с последействием, в силу чего выигравшие и проигравшие далее выступают как носители разного статуса, а охраняемые ими пространства — «разнокачественны» в смысле возможности противостоять попыткам захватов (рис.5—6). Это т. н. (+)-исход, в противоположность (—) или (0): в первом случае птицы теряют взаимный интерес и разлетаются, во втором – в их поведении ритуализованные демонстрации исчезают полностью, они начинают друг друга клевать и бить без какого-либо результата до полного изнеможения, иногда и серьёзных ранений. Оба исхода безуспешны в том смысле, что не происходит дифференциации поведенческих ролей, статуса индивидов и «качества» территорий. К трём этим исходам ведут качественные различия в поведении дятлов до их наступления, в организованном обмене демонстрациями и действиями (рис.8), почему все три выделяются объективно, это не интерпретация наблюдателя.
Сам территориальный конфликт больших пёстрых дятлов организуется своего рода «наращиванием ставок» ритуализированной агрессии (рис.8). Участники как бы соревнуются, cменяя демонстрации №№1–8 в направлении от менее ко всё более эффективным (хотя в данном случае главное, что — в одну сторону и «наперегонки»). Смена сопровождается «тычком» — выплеском прямых действий агрессии: клевков, выпадов, прыжков в местонахождение оппонента (т. н. вытеснение, англ. supplanting attack) или, наоборот, бегства — отступлений, отскоков, отворотов головы от оппонента.
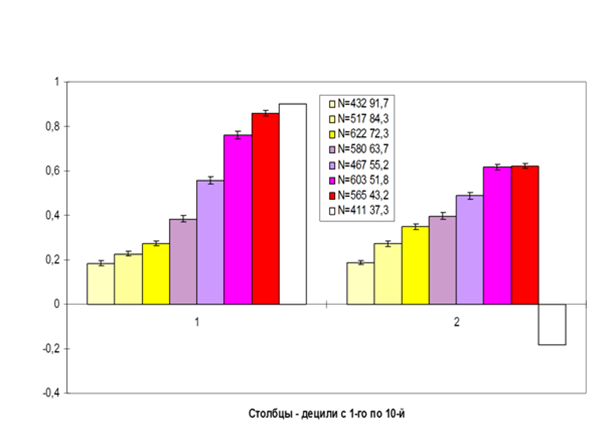
Рис.8. Динамика территориальных конфликтов больших пёстрых дятлов (охрана территорий в поселении «Лосиного острова», 24 птицы, сентябрь 1985–февраль 1986 г.г.). Обозначения. Ось абсцисс – время взаимодействия (использовали относительную оценку времени, в % до окончания процесса, с 1-го по 9-й децили), ордината – средняя эффективность демонстраций, адресованных оппоненту (о ней см.ниже), рассчитана по периодам, соответствующим децилям. 1 — динамика поведения будущего победителя, 2 – будущего проигравшего. Источник. «От стимула к символу», Ч.1.
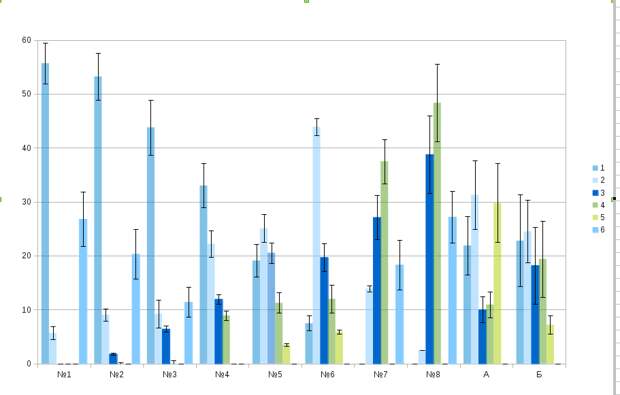
Рис.9. Состав «выплеска» прямых действий агрессии и/или бегства, сопровождающего демонстрации №№1-8. Обозначения. 1- Клевки, в т.ч. с подскоком/подлётом, 2 — Замирание на месте, прекращение локомоции, 3 — Отворачивания и отскоки от противника, 4 — Открытое бегство, в т.ч. с отлётом, 5 — Смещённые реакции, в т.ч. долбление как при кормодобывании, «сон» и пр., 6 — Вообще без сопровождения прямыми действиями. №№1-8 — демонстрации ритуализованной агрессии и бегства, специфичные для охраны территории, А — иные демонстрации: все прочие (преимущественно ухаживательные), встреченные во взаимодействиях охраны территории, патрулирования границ и/или «примешанные» к дистантным угрозам, Б — иные экспрессивные реакции: не входят в 100%, т.к. свободно сочетаются с действиями «выплеска» 1-6. Примечание. Соотношение трёх категорий противоположно направленных действий (атаки 1, бегства 3, 4, остановки и замирания, в т.ч. перед переходом к обмену демонстрациями на месте, без перемещений, клевков, отскоков и пр. 2) указывает на мотивационную подоснову предшествующих им демонстраций. Оно же, фиксируемое между демонстрациями (подобные паузы бывают весьма значительными), отражает конфликт побуждений к нападению и бегству у данной особи. Этим первые и вторые позволяют оценить мотивацию независимо от отслеживания обмена демонстрациями и его следствий — побед, поражений и иных исходов конфликтов, выступают для них внешним контролем.
Интенсивность «выплеска» прямых действий (количество движений и их экспрессия — скажем, атака в сравнении с выпадом, отскок в сравнении с отворотом корпуса и пр.) прямо пропорциональны «близкодействию» каждой из восьми демонстраций (рис.9, см. также Фридман, 2013б, разд.1.1, аналогичные данные по другим видам: Бутьев, Фридман, 2005). Соотношение действий разных типов (нападения, бегства и любых иных, не относящихся к агонистическим) в его составе соответствует мотивации демонстратора, отражая конфликт побуждений «внутри» него, почему наиболее вероятно предъявление этой, а не иной демонстрации (из восьми). Последнее обусловлено «конфликтом интересов» во внешнем мире участников — владельца участка и захватчика, оспаривающего у него большую или меньшую часть периферии (см. «От стимула к символу», Ч.2, табл.2). Т.е. 8 демонстраций территориальной агрессии выступают знаком-посредником, одновременно «указывающим» «конфликт интересов» между владельцами (интенсивность последнего — внешний референт знака) и выражающим конфликт побуждений атаки и бегства в каждом из них, но также в ритуализованной форме, «указывающей» на него символически, а не прямо, как «выплеск». Согласование обоих конфликтов достигается перемещением дятлов «вдоль» радиального «среза» участка (разного масштаба, от полётов-погонь в пространстве десятков метров до преследований в кроне одного дерева (метры), в зависимости от уровня возбуждения), пока не остановятся и перейдут к демонстрированию в точке, «уравнивающей» их интенсивности. См. «От стимула к символу», табл.10.
Так или иначе, каждый дятел пробует опередить второго в сменах демонстраций, сохранить постоянство их скорости и стереотипность демонстрирования, но одновременно «мешает» то же делать противнику, «обрушивая» на него именно в эти моменты действия выплеска, мешающие всему этому: удары, подскоки, да и отступления, побуждающие клюнуть в ответ. И всё это происходит тем сильней, чем более эффективная демонстрация им выбрана. Взаимные перемещения птиц, переплетения веток и т. д. общие обстоятельства, искажающие силуэт каждого демонстратора препятствуют разрешению первой задачи не меньше, чем активное противодействие их друг другу — как колеблемая ветром трава в агрессивных конфликтах агам A.muricatus (Peters, Evans, 2003ab, 2007, цит. по Фридман, 2013а, разд.3.9).
В какой-то момент один дятел распластывается, прекращая прямые действия в адрес противника — и клевки, и отскоки. Второй сразу же прекращает «тычки», но ещё 1—10 мин. удерживает прежнюю демонстрацию. Или, реже, клевки, выпады и отскоки ритуализируются: дятел останавливает клюв при ударе, «замахнувшись», отдёргивает или отворачивает голову, не пробуя реально поранить противника. Во втором случае отскок также превращается в отворот. Частота таких случаев возрастает в зависимости от плотности поселения с 2,4% в наиболее разреженных до 11,8% в самых плотных (F=3,04; df1=5; df2=2317; P<0,01, данные по 113 птицам за 9 сезонов). Влияние года и места наблюдений не значимо (F=1,4; P>0,05).
Это — конец взаимодействия: возобновлений конфликта после распластывания одной птицы и завершения «выплеска» другой не отмечали ни разу. Он наступает существенно после того, как один дятел стал устойчиво «опережать» в сменах демонстраций другого, т. е. после появления асимметрии в в их поведении проходит от 15 до 30% времени взаимодействий, включающих >1 смены демонстраций (в абсолютных значениях — от 20 с до нескольких минут). У больших пёстрых дятлов взаимодействие завершается много позже обнаружения явной асимметрии в демонстрировании, указывающей будущего победителя с высокой (и растущей во времени) вероятностью. То же фиксируется в коммуникации разных позвоночных, от рыб до млекопитающих и, видимо, универсально.
Первого дятла, не сохранившего избранную первоначально стратегию демонстрирования, сочли проигравшим, демонстрацию, удерживавшуюся перед распластыванием — «сигналом поражения». Каждая из единиц поведения благодаря близкодействию здесь обуславливает события «следующего шага процесса», а благодаря дальнодействию — общий исход, но опосредовано. Второго дятла, реализующего раз избранную стратегию до конца, сочли выигравшим, последнюю из предъявленных им демонстраций — обусловившей его выигрыш («сигнал победы»). Так можно определить разнотипные варианты исходов конфликта (победа или поражение, достигнутые употреблением 8 демонстраций разной эффективности) объективно, а не по впечатлениям наблюдателя. Принятое деление подтверждается реакцией «третьих» особей, в следующие полчаса-час пробующих захватывать периферию территории именно проигравшего (или маркировать там), и патрулировать (с маркировкой) границу своей территории, обращённую к выигравшему, но не наоборот. Эта асимметрия реагирования «зрителей» взаимодействия также отсутствует в случае (0)- или (-)-исхода конфликта. Сходным образом определяли выигравших и проигравших во взаимодействиях других контекстов, скажем, ухаживания (Фридман, 2013б, табл.2).
Наш анализ показывает, что эти три принципа:
1) «соревновательность» с «наращиванием ставки» ритуализированной агрессии до
2) прекращения сопротивления одной (проигравшей) особью и и прямых действий другой (выигравшей) конфликт, с
3) остановкой демонстрирования у обоих организуют агонистические взаимодействия везде, где это специально исследовали (см.выше).
И не только агонистические. Однотипно организованы взаимодействия прочих контекстов: ухаживание за потенциальным партнёром и/или самца с самки в уже образовавшейся паре (на этапе консолидации), умиротворения партнёра и/или доминанта и пр. Видимо, это универсальный способ организации взаимодействий: взаимно-дополнительные объяснения, почему так, одновременно даны и нами («От стимула к символу», Ч.2, табл.2), и М.Е.Гольцманом (2016).
Таким образом, (+)-исход означает дифференциацию на выигравшего и проигравшего участников взаимодействия, «тотипотентных» в его начале. С момента появления асимметрии в обмене демонстрациями определимо заранее, кто будет первым или вторым, и с тем большей вероятностью, чем больше прошло времени.
Выигрывает животное, «ведущее» в смене демонстраций на стадии «состязания», с тем большей вероятностью, чем устойчивей это «опережение» и скоординированней смены обоих. В результате моменты смен демонстраций (через некоторое время — и действий участников) «подгоняются» друг к другу всё точнее и точнее по ходу процесса; после выявления асимметрии временной «зазор», достигнув минимума, дальше не меняется (2-4 с., максимум до 8). «Отстающая» особь («ведомая» в сменах демонстраций и связанных с ними форм поведения, рис.8) особь почти гарантированно проигрывает — при сохранении «связанности» моментов смен после выявления асимметрии. Следовательно, (+)-исходы взаимодействий
а) «превращают» дифференциацию, ранее бывшую гибкой, ситуативной и изменяемой, в константные характеристики особей. В следующих взаимодействиях того же типа выигравший и проигравший дятлы с большей вероятностью будут соответственно активным и пассивным партнёрами, «ведущим» и «ведомым» в обмене демонстрациями, даже когда территориальный конфликт случается не между ними, а между одним из них и «третьей особью».
б) «закрепляют» её надолго: как минимум до следующего взаимодействия, по результатам которого она может быть «пересмотрена»,
в) делают её «явной» и «общепризнанной» для всех особей, «подключённых» к коммуникативной сети — как резидентов, так и «активных вселенцев», только пробующих занять территорию. Поскольку победа одной и поражение другой особи в конкурентном обмене демонстрациями и действиями достигнуты «по правилам», и специфическая организация последнего, что описана выше, на всём протяжении процесса реализована устойчиво и точно, без «сбоев». См. данные по американским чижам Spinis tristis (Popp, 1987c), молодым большим пёстрым дятлам при расселении (Фридман, 1996), крысам Rattus norvegicus (Lehner et al., 2011) и пр.видам.
На этой особенности (+)—исходов взаимодействий, агонистических и прочих (привлечения партнёра, ухаживания в паре, и т. д., Фридман, 2013, табл.10) основана эффективность «подглядывания». Именно оно связывает компаньонов в общеполезную «сеть», вместо того чтобы, как верят социобиологи, быть формой «обмана» (Фридман, 2017аб).
Напротив, учащение «сбоев» по ходу взаимодействия, нарушающих специфическую организацию обмена демонстрациями и действиями, ведёт к (0)— или (-)—исходу. В первом случае участники теряют интерес друг к другу, прекращают активность, и в конце концов разлетаются. Во втором картина внешне противоположна — они прекращают демонстрировать, взаимодействие «срывается» в беспорядочные погони и драки, часто с ранениями одной птицы или обеих; всё это продолжается десятки минут, иногда часы, т.ч. участники демонстрируют полное изнеможение. Оба «неположительных» исхода воспроизводимы экспериментально: подставлением чучела к местопребыванию владельца участка или зеркала — к территориальным парам в колонии, «неспособных» адекватно ответить на их демонстрации, вызванные такой имитацией «захвата» (Фридман, 2013б, разд.2.9).
При (0) или (-)-исходах взаимодействия безуспешны. Дятлы (и прочие виды в данной ситуации) щедро тратят на них время и силы, сильно рискуют стать жертвой хищника и/или получить травм
Свежие комментарии