Ещё одна работа над ошибками — что не так с концепцией «самодоместикации» бонобо. Я сперва ей доверял, т. к. поскольку она настолько распространена среди исследователей доместикации, что приводится как «давно известное», без сравнения с конкурирующими объяснениями, см. взгляд на антропогенез Л.Н.Трут (2008), ведущего автора из продолжателей беляевских опытов с одомашниванием лисиц.
***
Когда в руках у тебя молоток, любая проблема кажется гвоздём. Так произошло с концепцией доместикации Д.К. Беляева (1983): отбор на дружелюбное отношение к человеку, вольеру, вообще контролируемой им обстановке вокруг животного вместо прежней агрессии или тревожно оборонительной реакции меняет механику развития (задержка миграции клеток нервного гребня), она тянет за собой изменений морфологии, гормонального состояния, и поведения в отношении хозяев, окружения, нового-неизвестного и т. д.
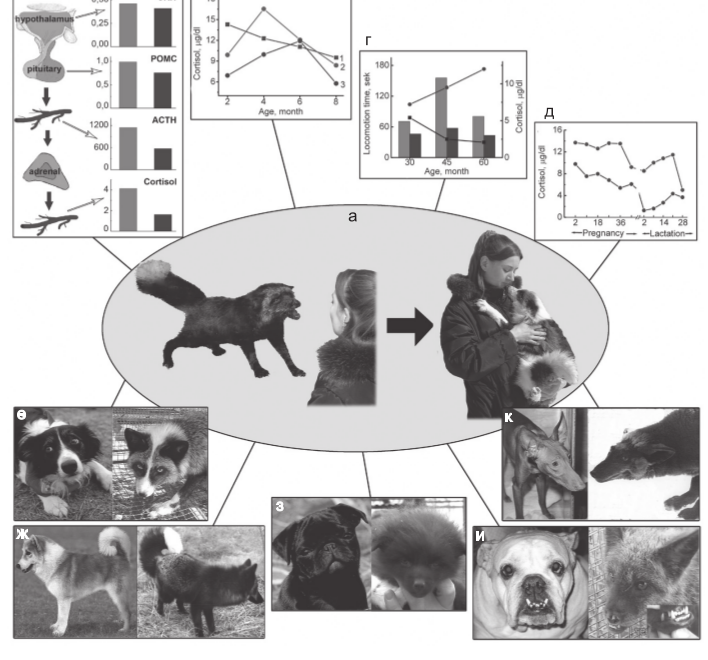
Рис.1. Схематическое представление основных результатов эксперимента Д.К.Беляева.
Обозначения. а – трансформация поведения лисиц в ходе отбора: слева – лисица из неселекционируемой по поведению промышленной популяции. Агрессивная реакция на человека наиболее типична для этих лисиц: справа – лисица из экспериментальной доместицируемой популяции. Поведение, характерное для собак, сформировалось у этих лисиц в результате длительного отбора;
б–д – показатели функциональной активности гипоталамо-гипофизарно-надпочечниковой системы (ГГНС);
б – экспрессия гена (или уровень мРНК) кортикотропин-рилизинг гормона (CRH) в гипоталамусе и пропиомеланокортина (POMC) в гипофизе; уровни адренокортикотропного гормона (АКТГ) и кортизола в плазме крови у неселекционируемых (слева) и ручных (справа) серебристо-черных лисиц;
в – возрастные изменения уровня кортизола в плазме крови у неселекционируемых и ручных серебристо-черных лисиц. 1, 2 – неселекционируемые лисицы: 1 – проявляющие агрессивную реакцию на человека, 2 – не проявляющие ее; 3 – ручные лисицы с высокой оценкой доместикации;
г – суммарное время двигательной активности – индикатора исследовательского поведения и уровень кортизола в плазме крови у неселекционируемых и ручных серебристо-черных лисиц в возрасте 1–2 месяца: двигательная активность изображена графически; нижняя линия – неселекционируемые, верхняя – ручные; уровень кортизола – в виде гистограммы (левые столбики – неселекционируемые, правые – ручные);
д – уровень кортизола в плазме крови у серебристо-черных лисиц при беременности и лактации: (верхняя линия – неселекционируемые, нижняя – ручные);
е–к – некоторые морфологические изменения, имеющиеся у собак и возникшие у лисиц доместицируемой популяции; е – сходный характер депигментации волосяного покрова у собак и лисиц: слева – собака бордер-колли; справа – ручная лисица; ж – манера держать хвост свернутым в кольцо: слева – исландская собака; справа – ручная серебристо-черная лисица; з – у отдельных ручных лисят повисают уши и укорачивается лицевой череп, что характерно для некоторых пород собак: слева – собака мопс; справа – ручной лисенок; к – удлинение лицевого черепа у некоторых пород собак и у ручных лисиц: слева – фараонова собака; справа – лисица; и – относительное удлинение нижней челюсти (перекус), наблюдаемое у английского бульдога, происходит и у некоторых ручных лисиц: слева – английский бульдог; справа – ручная лисица.
Источник. Трут Л.Н., 2008. Эволюционные идеи Д.К. Беляева как концептуальный мост между биологией, социологией и медициной// вестник вогис. Т12. №1/2. С.7-19.
Когда она стала модной на Западе, её стали прилагать ко всему, хоть в чём-то сходному с данным процессом, и как правило, неудачно. Скажем, к урбанизации «диких» видов птиц и млекопитающих: что здесь не так, см. «Урбанизация — не доместикация».
Домовые мыши в амбаре: здесь самодоместикация более-менее подходит
Содержание
Дальше идею «самодоместикации» применили к изменениям, отмеченным у домовых мышей Mus musculus из Швейцарии, живших в амбаре, недоступном для хищника (Geiger et al., 2018), бонобо в сравнении с обыкновенным шимпанзе (Hare et al., 2012), лис, освоивших города Англии и не только (Parsons et al., 2020, см. пересказ по-русски). Мол, всякое понижение риска хищничества и/или угрозы агрессии вызывает отбор на миролюбие с его гормональными и морфологическими коррелятами, и самые явственные среди них — пегость, вислоухость, укорочение лицевой части черепа и относительное уменьшение мозга.
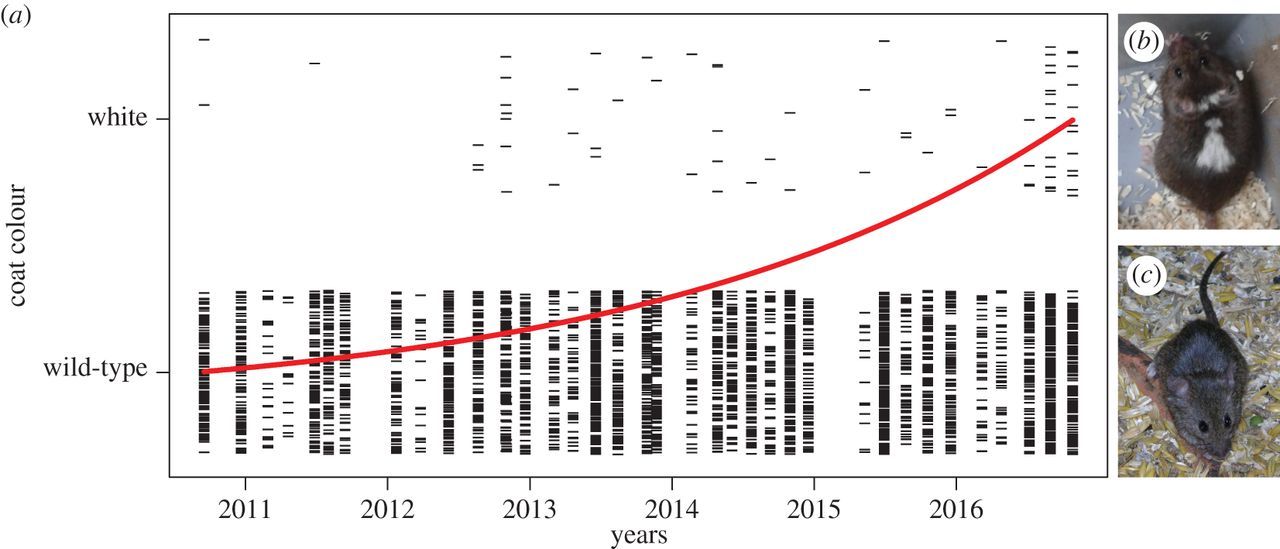
Рис.2. Направленное развитие пегостей у мышей из амбара в сравнении с мышами «дикого типа», Geiger et al., 2018.
С мышами это объяснение более-менее проходит. У домовых мышей, изолированных в амбаре от хищников, за 2002-2016 гг. появились типичные доместикационные изменения — пегости, укорочение лицевой части черепа, уменьшение абсолютных и относительных длин головы и тела (Geiger et al., 2018, см.рис.2-3).
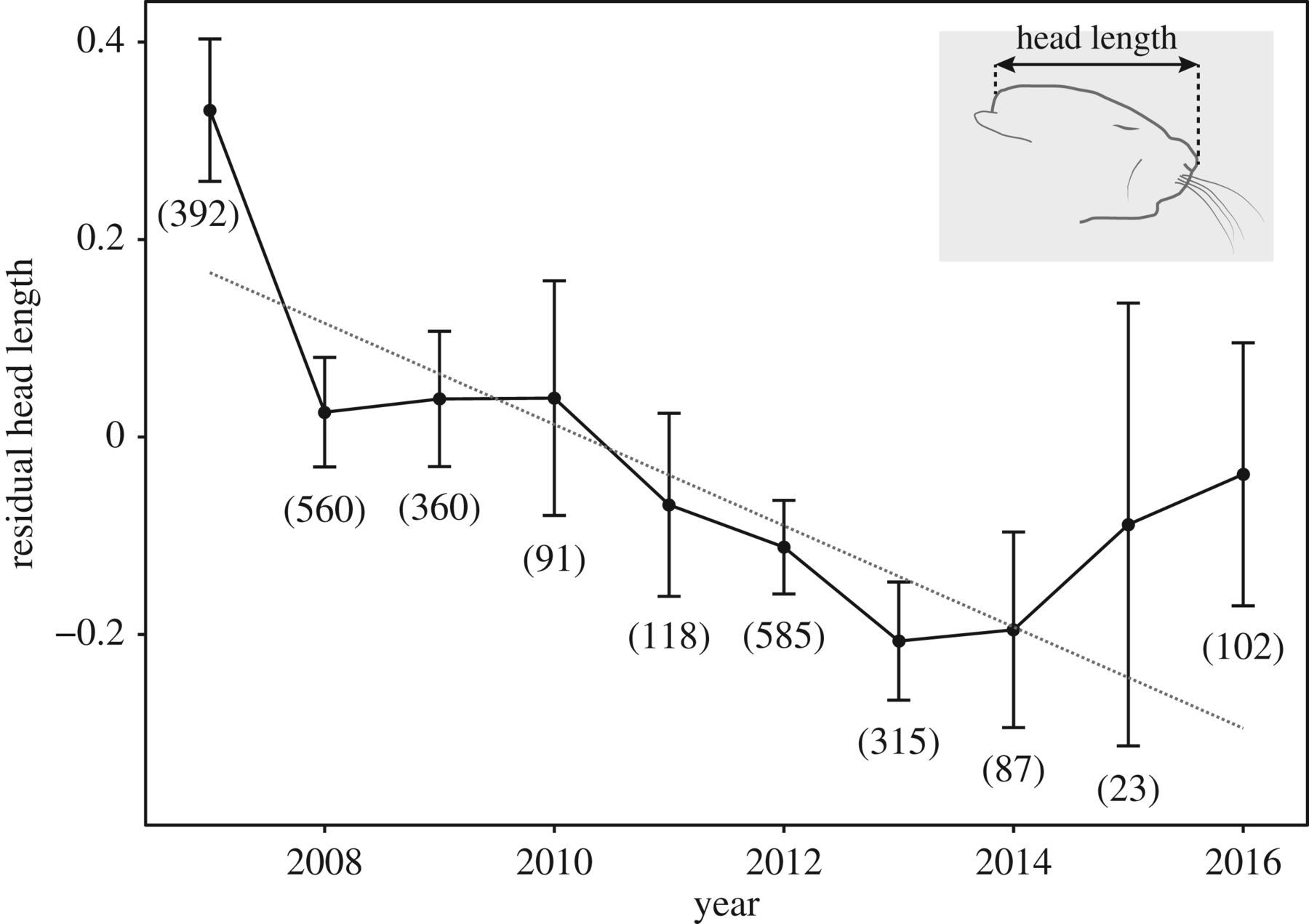
Рис.3. Направленные изменения лицевой части черепа у домовых мышей в амбаре, Geiger et al., 2018.
Скорость направленного сдвига морфологии, исчисленная, как обычно делается для микроэволюции, в дарвинах и холдейнах (табл.1), лежит в тех же пределах что элементарные эволюционные явления, меняющие признаки черепа в других популяциях «диких» видов грызунов (всего 8), но значимо выше — сокращение длины головы соответствует 4017 дарвинов и 0.499 холдейнов, тест Вилкоксона V = 0, p = 0.0078. Это отражает общее ускорение микроэволюции, когда её вызывает антропогенная трансформация ландшафта, и особенно урбанизация (здесь эволюция идёт быстрей всего).
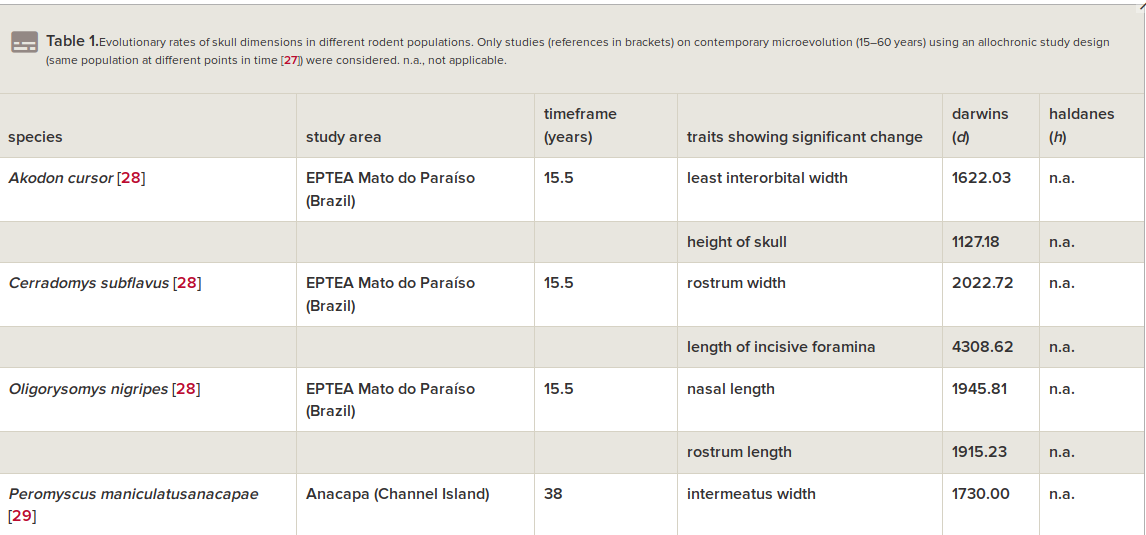
Табл.1. Скорости микроэволюционных изменений черепа в «диких» популяциях грызунов, взятые для сравнения. Из: Geiger et al., 2018
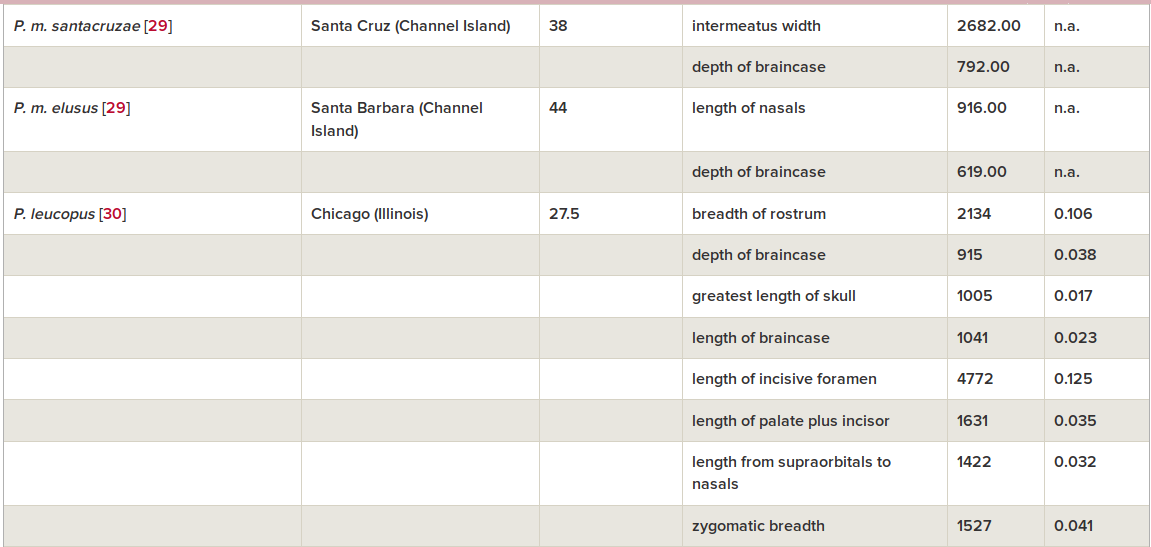
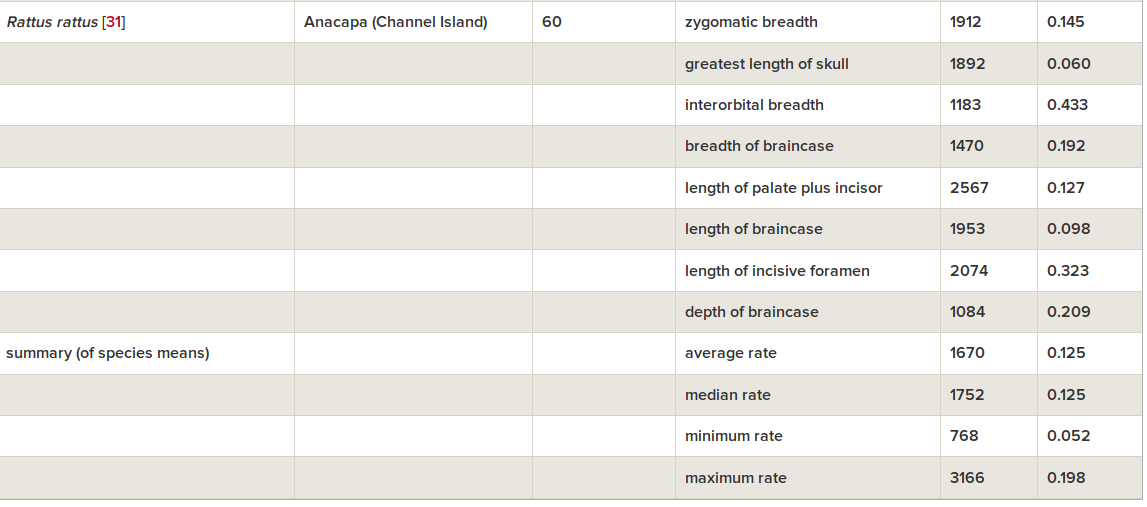 Авторы полагают, что, отлавливая и измеряя мышей, непреднамеренно осуществили отбор; так бывает с другими искусственно разводимыми видами (Артамонова, Махров, 2006). Из-за этого беспокойства антропофобные мыши рассеивались из амбара, оставались и размножались там лишь самые ручные, а его недоступность для хищников усиливала эффект. Правда, не были исключены альтернативные объяснения происходящего: их предшествующая синантропизация. Мыши уже были комменсалами при заселении Европы, в т.ч. менее агрессивными к сородичам, с ослабленной регуляцией «запуска» тревожно-оборонительных реакций, как и доместицированные виды. Происходящее также можно объяснить дрейфом генов, а преобразования черепа — сменой состава корма сравнительно с популяцией-источником (просыпавшееся зерно и фураж, Geiger et al., 2018).
Авторы полагают, что, отлавливая и измеряя мышей, непреднамеренно осуществили отбор; так бывает с другими искусственно разводимыми видами (Артамонова, Махров, 2006). Из-за этого беспокойства антропофобные мыши рассеивались из амбара, оставались и размножались там лишь самые ручные, а его недоступность для хищников усиливала эффект. Правда, не были исключены альтернативные объяснения происходящего: их предшествующая синантропизация. Мыши уже были комменсалами при заселении Европы, в т.ч. менее агрессивными к сородичам, с ослабленной регуляцией «запуска» тревожно-оборонительных реакций, как и доместицированные виды. Происходящее также можно объяснить дрейфом генов, а преобразования черепа — сменой состава корма сравнительно с популяцией-источником (просыпавшееся зерно и фураж, Geiger et al., 2018).
С бонобо «самодоместикация» совсем не проходит
А вот для объяснения отличий бонобо от обыкновенного шимпанзе (как делается Брайаном Хейром и Ванессой Вудс, см. Hare et al., 2012 или «Почему собаки гораздо умней, чем вы думаете») «доместикация» не годится вовсе, даже «само». См. связанные между собой различия домашних животных от их диких сородичей по морфологии, физиологии, поведению из табл.1 в статье Hare et al. (2012).
Табл.1. Доказательство присутствия доместикационного синдрома у разных видов
| Бонобо — обыкновенный шимпанзе | Домашняя собака — волк | Экспериментальные лисицы Беляева — контроль | Домашние морские свинки — дикие | |
| Агрессия | Снижение интенсивности внутри- и межгрупповой агрессии у бонобо | Снижение интенсивности внутри- и межгрупповой агрессии у собак | Первые, но не вторые неагрессивны к людям | Первые, как и лабораторные крысы и мыши, гораздо менее агрессивны к сородичам |
| Физиология | Бонобо уязвимей к стрессу, особенно в конкурентной ситуации, поведение совладания при стрессе у них более пассивное, самки бонобо характеризуются скрытой овуляцией и рецептивны большую часть цикла | Собаки ослабляют действие стресса, взаимодействуя с людьми; множественные периоды половой рецептивности у самок собак, но не волков. | Повышенный уровень базального серотонина и сниженный — кортикостероидов у первых в сравнении со вторыми, у них же — множественные периоды сексуальной рецептивности, | Домашние свинки приобретают пегости, объём черепа у них уменьшен, Воздействие стресса на первых в сравнении со вторыми ослаблено в присутствии конспецификов, взаимодействии с ними и при исследовательской активности. |
| Морфология | Общая ювенализация, в т.ч. тимус работает даже у взрослых особей. Объём мозговой коробки уменьшен, ниже половой диморфизм по клыкам, депигментация губ и хвоста (но не пегости). | Собаки в сравнении с волком обладают уменьшенными мозговой коробкой, половым диморфизмом по крыкам, появляются пегости и вислоухость. | снижение полового диморфизма, в т.ч. по развитию хищных зубов, пегости, феминизация черепа. | У момашних свинок — уменьшенный объём черепной коробки, пегости |
| Просоциальное поведение | Во взрослом состоянии больше играют, большую роль в социальных отношениях занимает секс, а не агрессия, сравнительно с обыкновенным шимпанзе. Они добровольно делятся пищей и терпимы к присутствию других при еде | Первые проявляют больше игрового и социосексуального поведения в зрелом возрасте, их больше привлекают люди, чем конспецифики | Первые в зрелом возрасте больше заинтересованы в привлечении людей и взаимодействии с ними | Домашние свинки более склонны играть и демонстрируют больше полового поведения; лабораторные грызуны более спокойны при взятии в руки. |
| Задержка развития (ювенализация) | Задержанное развитие социального поведения и познания относительно шимпанзе (ювенализация). Бонобо более зависимы от матери, чем шимпанзе; медленнее развивают социальные навыки в задачах. связанных с добычей пищи и конкуренцией за кормление. | Задержана выдача взрослой тревожно-оборонительной реакции у собак; во взрослом возрасте сохраняются ювенильная реакция страха и средства коммуникации характерные для щенков | У первых задержана выдача взрослой реакции страха; щенячьи средства коммуникации сохраняются и у взрослых | Меньшая агрессивность экспериментальных грызунов — следствие задержки развития. Взрослые сохраняют ювенильный коммуникативный репертуар |
| Когнитивные способности | Бонобо чувствительней к «социальным подсказкам» людей и лучше справляются с задачами, требующими социальной терпимости | Собаки чувствительней к «социальным подсказкам» людей | Экспериментальные лисы чувствительней к «социальным подсказкам» людей | Домашние свинки лучше диких справляются с тестом на пространственное |
Примечания. У самцов бонобо в конкурентной ситуации (во всех или как минимум в большинстве) повышается уровень кортизола, т.е. конкуренция их стрессирует, им плохо. У большого шимпанзе, напротив, повышается уровень тестостерона, т.е. у них кураж, увеличивающий силы. У людей, в общем, как у бонобо, но если конкурентный мотив осознаётся как «лично значимый», до конкуренции есть сильное желание победить — как у шимпанзе. Так действует социальное влияние, я считаю, поскольку всякие представления и желания «загружаются» внутрь реакциями других людей на нашу активность.
В ней всё хорошо соответствует модели доместикации, лишь одна пара: «бонобо — обыкновенный шимпанзе» здесь лишняя; увы, статья обосновывает как раз самодоместикацию P.paniscus — и ошибается. Авторы доказывают, что различиях между видами шимпанзе того же рода, что отличия домашней собаки от волка, домашней морской свинки — от дикой, лабораторных крыс и мышей — от пасюков и «диких» Mus musculus/M.domesticus, беляевских, доброжелательных к нам лис, — от диких родичей.
Что справедливо везде, кроме восприятия «социальных подсказок» со стороны людей и сородичей при решении когнитивных задач (полужирный курсив в табл.1). Оба вида шимпанзе равно не обращают на них внимания, не ищут их и т.д., притом что существа весьма умные. Авторы утверждают обратное, однако работы, на которые они ссылаются (Herrmann et al., 2010; Hare, Kwetuenda, 2010; Wobber et al., 2010) совсем о другом: большей социальной связанности и социальной зависимости бонобо во всех контекстах взаимодействия. Pan troglodytes «бескорыстно» (вне контекста доминирования или возможности спаривания) делятся пищей с друзьями внутри «своей» группы, то бонобо — также с чужаками и с членами других групп (Gylby et al., 2010, цит.по обзору в: Fruth, Hohmann, 2018).
В решении же когнитивных задач оба вида равно нечувствительны к «подсказкам», мало способны к совместному действию. И, главное, в понимании и использовании коммуникативных жестов бонобо не превосходит шимпанзе в той же степени, в какой собака — волка и другие пары в этой таблице (Herrmann et al., 2010, table 1; см.также коммуникативное использование жестов обоими видами). Новые данные наблюдений за дикими бонобо показали регулярное использование орудий и не меньшее развитие хищничества, чем у «больших» шимпанзе. Они нивелировали прежде утверждавшееся отличие от P.troglodytes: мол, бонобо лучше решает задачи, относящиеся к «теории сознания» и причинности в социальных взаимодействиях, тогда как шимпанзе — к причинности физического мира и в инструментальной деятельности (см. «Близость к человеку: шимпанзе vs бонобо»).
Сегодня понятно, что у обоих видов мир социальных связей и физических явлений не отделён друг от друга, они равно способны строить планы и достигать результатов в том и другом, используя орудия или других особей, но «своими умом», без «подсказок» извне (см. обзор опытов М.Томазелло в: Федорович, 2015). Скажем, Girard-Buttoz et al. (2020) исследовали способность бонобо и шимпанзе к кооперации в предупреждении об опасности, предъявляя смертельную для них габонскую гадюку. Оба вида показали развитую «теорию сознания» (the theory of mind): «понимали», видит собрат змею или нет, и кричали лишь для невидящих (или же прекращали кричать, когда другая обезьяна увидела).
Однако у шимпанзе предупреждение развито лучше: обезьяны, прибывшие позже к змее, были меньше удивлены и испуганы, т. е. скорей всего, были предупреждены ранее, слышали крики, пока добирались и пр. Иными словами, шимпанзе «при прочих равных» более мотивированы сотрудничать внутри группы при предупреждении об опасности что хорошо сочетается с использованием агрессии для контроля пространства (Girard-Buttoz et al., 2020). Бонобо для той же цели используют дружественные взаимодействия: груминг, игру и секс, почему более склонны сотрудничать между группами. Недавно был описан первый случай дележа с «чужаками» мясом добытого дукера (Fruth, Hohmann, 2018).
Другой важный момент: бонобо архаичней Pan troglodytes по морфологии, особенностям онтогенеза, «конструкции» социальной системы. Первый ближе к общему предку рода Pan и «ветви», ведущей к австралопитекам и людям, второй — вид более продвинутый и молодой. Но во всех прочих парах в таблице доместицированный вид — потомок «дикого», не наоборот!
Анцестральность социальной организации бонобо
Архаичность социальной организации бонобо доказывается ещё и тем, что при определённых условиях а) взаимоотношения полов и б) территориальность обыкновенных шимпанзе повторяют вариант характерный для бонобо, притом что «в среднем» резко отличны от них. Первое см. «Механизм доминирования самок у бонобо», второе недавно изучено у западного подвида обыкновенного шимпанзе в лесу Тай.
За 20 лет наблюдений за взаимоотношениями между соседними группами и социальной динамикой внутри них в лесу Тай было показано, что действия самцов важны для расширения территории, но её сохранение и одержание побед над соседями, пробующими что-то с краёв отобрать, или просто хотят пройти через их участок, зависит от самок — насколько они поддерживают самцов, насколько тесны социальные связи в данной группировке (Lemoine et al., 2020). В целом, чем выше социальная связанность и социальная зависимость между особями (в т.ч. доминантов от подчинённых им самок), тем прочнее она закрепляется в определённом пространстве.
Поэтому размеры и/или сплочённость группы важней для победы в стычках, чем количество «бойцов» (в целом возрастающее с увеличением территории). Чем они выше, тем — при прочих равных — выше размер групповой территории и слабей прессинг соседей. Одновременно уменьшается риск поранения и/или гибели в собственно стычках, в сравнении с восточноафриканской ситуацией, когда одни самцы во время рейдов противостоят других (успех рейда ведёт к поранению и даже гибели «чужих» особей обоих полов). Т.е. здесь, как и в наблюдениях де Вааля за шимпанзе в зоопарке Арнема, видим переход к «бескровному» варианту охраны пространства, типичному для бонобо.
Дело в том что второй вид не менее территориален, чем первый, в том смысле, что группы, обменивающиеся особями и управляемые матриархами, привязаны к определённым территориям, их границы отлично известны и строго очерчены, это пространство регулярно обследуют, высматривая «чужаков» из соседних групп, которых вовнутрь не пускают. В отличие от обыкновенного шимпанзе, их останавливают без агрессии и убийств, а вовлечением на границе в дружелюбные взаимодействия: игры, груминг, сексуальные контакты (Hare, 2012). Агрессивные проявления отмечаются как часть этих взаимодействий, но прямая агрессия и травмы редки, и сильно варьируют между сообществами, в одних единичны, в других — до трети случаев встреч. Будучи наиболее обычны в начале контактов, они сильно сокращаются по мере знакомства обеих групп, замещаясь грумингом, сексом, совместным кормлением и т.д. позитивом. Недавно стало известно, что при таких встречах бонобо также делятся с «соседями» добытым в этот момент мясом, что немыслимо для шимпанзе (Fruth, Hohmann, 2018).

Рис.4. Всматриваясь в кусок мяса, бонобо собираются вокруг обладателя ноги дукера
Сегодня известно, что бонобо не менее «мясоядны», чем шимпанзе, и в природе регулярно охотятся, в первую очередь на лесных антилоп-дукеров Cephalophus callipygus, С.wayni, но также мелких обезьян, птиц и даманов, см. Wakefild et al., 2019; обзор с наблюдениями Fruth, Hohmann, 2002, 2018. Их мясом они делятся друг с другом точно также (рис.4) как и фруктами, что описывали ранее (рис.5; просящие бонобо собрались вокруг самца с плодом хлебного дерева Trecula africana).

Рис.5.
В связи с особенностью социальной системы бонобо делёжи чаще всего контролируются взрослыми самками (притом что самцы как у всех понгид, крупнее и сильнее самок). Нечасто, но регулярно проявляют агрессию в отношении тех, кто пробует поесть мяса самостоятельно, особенно взрослых самцов, но делятся с теми, кто выпрашивает, особенно молодыми самцами. А слишком назойливым просителям могут сжатым кулаком и по морде дать (Wakefield et al., 2019).
Независимо от размера куска (мяса, плода, мякоти сердцевины или листьев) и как он добыт — группой или в одиночку, имеют место выпрашивание корма другими, отрывание и передача кусков (рис.4-5), чем укрепляются узы между особями в сообществе. Отсюда можно предположить, что в линии к человеку способность к делёжке и общее нам всем стремление к равенство возникли и эволюционировали в связи с добычей, а потом производством любой пищи, не только мясной посредством охоты, как считалось ранее, добыча, важная для сплочения сообществ, включала всех особей, а не только охотящихся, в первую очередь самок с подростками.
Вместо этого [для понимания человеческой эволюции] важно рассмотреть, может ли определённая пища монополизироваться, и как это влияет на социальные связи между вовлечёнными [в её добычу самками]. Вместо того, чтобы считать энергетически ценное мясо пейсмекером политических отношений у гоминид, полезней анализировать процессы, благодаря которым корм, в принципе вызывающий конкуренцию и легко монополизируемый, тем не менее бескорыстно делится с другими. В частности, эти процессы могут быть независимы от величины и энергетической ценности рассматриваемых продуктов (Fruth, Hohmann, 2018).
У обыкновенного шимпанзе дележи стимулируют выделение окситоцина, играющего ключевую роль в укреплении социальных связей и сотрудничестве. Для орангов и бонобо показано, что пища выпрашивается не только ради неё самой, это механизм социального контроля. Так просители «проверяют» дружелюбность к ним высокоранговых особей, готовность защитить, оказать покровительство и в иных ситуациях, что, в свою очередь, влияет на поддержку со стороны подчинённых в «спорах» за высший ранг (почему выпрашивание и не зависит от ценности пищи).
Деление мясом дукера с чужаками впервые отмечено во встрече двух сообществ, западного (З) и восточного (В, состав их см.Fruth, Hohmann, 2018, table 1), в LuiKotale, Конго. Особи З полностью привыкли к наблюдателю и естественно ведут себя с 2007 г., В — с 2015 г. Все бонобо распознавались индивидуально, родство внутри З контролировалось по молекулярным маркерам. Отслеживание поведения и картирование местоположения особей по GPS вели ежедневно.
21 июня подгруппы В и З, т. н. партии, из 10 половозрелых самок и 5 самцов (подростки не учитываются) vs 12 самок / 3 самца, прослеживаемые наблюдателями, остановились у р.Бомпуза, сделали ночёвочные гнёзда. На следующий день они смешались в лесу западнее этой реки. В 14.04 Камилло, альфа-самец сообщества З, поймал дукера, и его сразу же обступили представители обоих сообществ. В 14.11 Камилло двинулся в крону высокого дерева, за ним следовали 9 самок с потомством. В следующие 30 минут эти самки (Gwen, Iris, Uma, Rio, Katie из З и Evimba, Agbaya, Kimya, Lombe из В) выпрашивали части дукера, которыми владел Камилло. Выпрашивание включало в себя пристальный взгляд, попытка обратить на себя внимание, протягивание рук, но не агрессия или попытка взять самому. Передача пищи от самца к самкам, как и в других случаях, была пассивной — они брали после «разрешения» взглядом, а не он давал своей рукой.
За это время 5 самок из З и одна из В (Agbaya) получили небольшие кусочки мяса и немедленно сожрали их. В 14.36 Agbaya оторвала всю голову дукера и разделила её между самками обоих общин и своим потомством. Iris получила ногу от туши, принадлежащей Камилло, и поделилась мясом со взрослыми самками своего сообщества, и своим потомством. Регистрации поведения обезьян за время дележа включали многочисленные потирания гениталиями между взрослыми самками из разных сообществ, как и внутри одного сообщества. Отмечено одно спаривание взрослого самца из В с самкой из З, и взаимный груминг между самками из разных сообществ.
В 15.02 туша перешла от Камилло к самке, не распознанной индивидуально. Прослеживание стало затруднённым из-за листвы и дождя, видимость ещё больше ухудшилась, когда обезьяны ушли в крону. Дальше они совсем стали невидимыми. Члены З не передвигались из этого места, В — были встречены позже в месте прошлой ночёвки, где построили гнёзда в период с 16.38 до 17.11.
Несмотря на присутствие многих других самцов, в дележе не участвовал никто, кроме Камилло. Никаких актов агрессии не отмечено ни между полами, ни внутри каждого из них, хотя такое регулярно бывает в других встречах данных сообществ. Чтобы поставить место дележа мяса с чужаками в контекст использования пространства В и З, на рис.6а и 6б приведёны данные от их местонахождении до (с 1 по 22 января) и после (22.01-13.02). По этим точкам строятся участки обитания как минимальный многоугольник, включающий все регистрации группы.
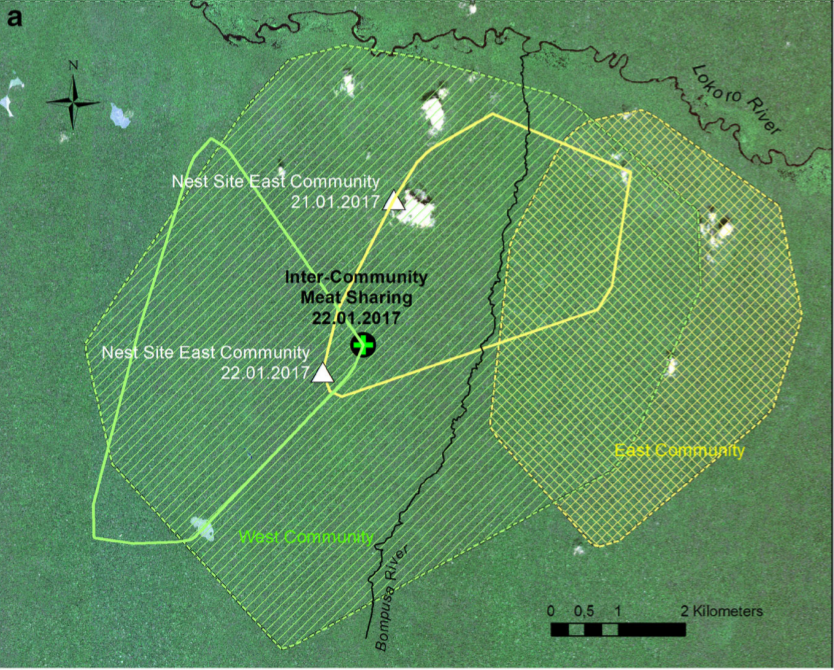
Рис.6а. Треугольники — места ночлега В и З, крест — делёж мяса дукера между ними
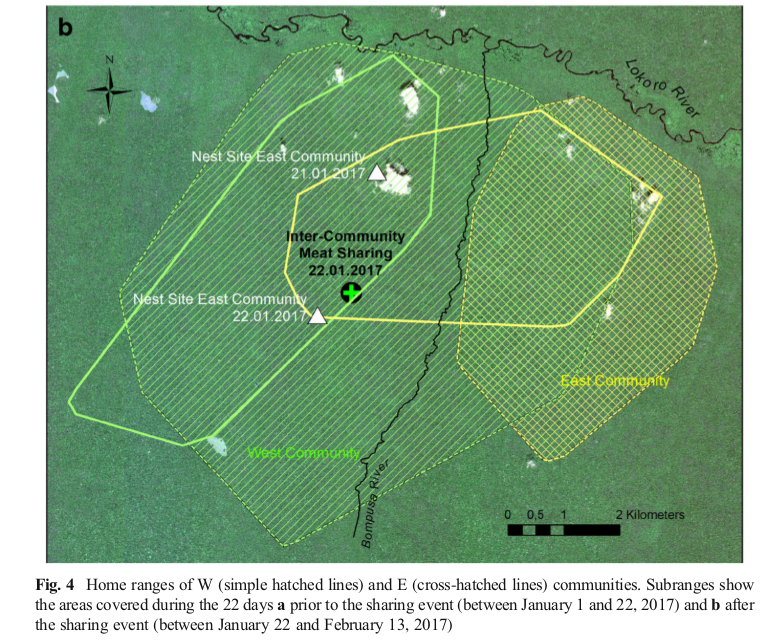
Рис.6б. Треугольники — места ночлега В и З, крест — делёж мяса дукера между ними, линии — границы индивидуальных участков.
Особи В находились восточнее р.Бомпузы (заштриховано в клеточку), участок обитания З значительно больше, но месяц перед делёжкой группа находилась в основном на существенно меньшей площади, в юго-западной части, а к середине января вернулись в его центр (простая штриховка). Особи В в этом период сильно углубились внутрь участка обитания З. Несмотря на в целом позитивный характер взаимодействия, в последующие дни они возвращались на свой участок, а не оставались на чужой территории, т. е. удержание пространства «заласкиванием» соседей работает.
Другой важный момент — происходящее не объяснить из социобиологической схемы, упирающей на наибольшую скрытость овуляции у бонобо, определяющей важность охрана самцами самок, длительных связей с ними, самочьего выбора «лучших» партнёров, в т.ч. как охотников (отсюда весьма популярная, но не работающая гипотеза «обмена еды на секс») в рамках которого увеличивается терпимость самцов к самкам. Увы, индивидуальные самцы бонобо при содержании с неволе ведут себя вполне деспотически, лишь в сообществе это сдерживается коллективными усилиями самок, «обмен» же «мяса на секс» опровергается данными.
Описанное у бонобо сходно с позитивными контактами на границе территорий разных групп охотников-собирателей, в т.ч. и довольно воинственных, вроде описанных ниже племён австралийских аборигенов. Эти контакты дружелюбны, чужим дарят понравившиеся им вещи, и спариваются кто с кем хочет. Из монетизации подобных обменов в предклассовом и классовом обществе родилась проституция, сперва храмовая, потом рыночная:
«…перенесемся на другой край Земли – в Западный Арнемленд, в Австралию. Живущий здесь народ гунвингу известен тем, что проводит со своими соседями ритуалы церемониального обмена под называние дзамалаг. Здесь угроза применения насилия выражена намного слабее – отчасти потому, что ситуация облегчается наличием системы кровнородственных связей, которая охватывает весь регион: никому не дозволяется жениться на представительницах своей родственной секции или заниматься с ними сексом вне зависимости от того, откуда они. Но любой человек из другой секции может быть потенциальным партнером.
Таким образом, для мужчины даже в общинах, проживающих далеко от его собственной, контакты с одной половиной женщин строго запрещены, а с другой – допустимы. Регион также объединяет местная специализация: у каждого племени есть свой товар, которым оно обменивается с другими племенами.
Ниже я привожу выдержку из описания дзамалага, которое в 1940‑х годах дал антрополог Рональд Берндт.«Здесь обмен тоже начинается тогда, когда, после первоначальных переговоров, чужаков приглашают на главную стоянку хозяев. В данном конкретном случае гости славились своими «ценными зазубренными копьями», а хозяева располагали хорошей европейской одеждой. Сначала группа гостей, состоящая из мужчин и женщин, вступает на танцевальную площадку стоянки, или в «круг», и трое из них начинают развлекать хозяев музыкой. Двое мужчин поют, а третий аккомпанирует им на диджериду. Вскоре приходят женщины принимающей стороны и нападают на музыкантов:
Мужчины и женщины встают и начинают танцевать. Дзамалаг завязывается тогда, когда две женщины гунвингу из родственной секции, противоположной той, к которой относятся певцы, «дают им дзамалаг»: они дарят каждому мужчине какой‑нибудь предмет одежды, ударяют его или касаются и увлекают на землю, называя его своим дзамалаг‑мужем и заигрывая с ним. Затем другая женщина из секции, противоположной той, к которой принадлежит мужчина, играющий на трубе, дает ему одежду, ударяет и заигрывает с ним.
Это кладет начало дзамалагу. Гости сидят спокойно, пока женщины из противоположной родственной секции подходят к ним, дают одежду, ударяют их и склоняют к соитию; в обстановке веселья, подбадриваемые аплодисментами, они позволяют себе любые вольности с мужчинами, пока продолжаются пение и танцы.
Женщины пытаются развязать набедренные повязки мужчин или коснуться их пениса и увести их за пределы «круга», чтобы совокупиться с ними. Мужчины, делая вид, что сопротивляются, удаляются со своими дзамалаг‑партнершами и совокупляются с ними в кустах подальше от огней, что освещают танцоров. Они могут дать женщинам табак или бусы. Когда женщины возвращаются, они отдают часть этого табака своим мужьям, которые побуждали их принять участие в дзамалаге. Мужчины, в свою очередь, расплачиваются табаком со своими дзамалаг‑партнершами.
Выходят новые певцы и музыканты, на них снова набрасываются, а потом увлекают в кусты; мужчины призывают своих жен «не скромничать» и подтвердить репутацию гунвингу как радушных хозяев; они и сами проявляют инициативу по отношению к женам гостей, предлагают им одежду, ударяют их и уводят в кусты. Бусы и табак переходят из рук в руки. Наконец, когда все участники совокупились хотя бы по одному разу и гости остались довольны приобретенной одеждой, женщины прекращают танец и встают в два ряда; гости выстраиваются в линию, чтобы отплатить им.
Затем мужчины‑гости из одной родственной секции танцуют перед женщинами из противоположной секции, чтобы «дать им дзамалаг». Они держат наперевес копья с лопатообразными наконечниками и делают вид, что вонзают их в женщин, но на деле лишь ударяют их наконечниками плашмя. «Мы не будем вас пронзать, ибо мы уже пронзили вас своими пенисами». Они дарят копья женщинам. Затем мужчины‑гости из другой родственной секции проделывают те же действия с женщинами из противоположной им секции и дают им копья с зазубренными наконечниками. Этим церемония завершается, после чего следуют щедрая раздача еды»
Давид Гребер. Долг. Первые 5000 лет истории.
В «экономических» экспериментах показано, что люди неклассовых обществ — бушмены !кунг, папуасы эйпо, — мотивированы делиться с «чужими» не менее, чем со своими, в отличие от земледельцев из классовых обществ и современных западных обывателей. В сходно построенных экспериментах в неволе то же делают оранги и бонобо, но не большие шимпанзе, и было показано что смысл дележа не в самой пище, а в укреплении просоциальной мотивации у обоих и повышения терпимости между чужаком и «хозяином» (из обзора в: Fruth, Hohmann, 2018).
С 2015 г. сообщества В и З встречались редко, дважды в год, но с начала 2017 г. встречи участились до в среднем 1 раза в месяц. Т.е. они были друг другу чужими; однако, т. к. у бонобо расселяются самки, в одной группе могли быть особи из другой, поселившиеся до 2007 г., т. е. отчасти «свои». Январский делёж мяса важен тем, что показывает новое измерение взаимной терпимости у бонобо, ранее неизвестное для них. Взрослый самец делил добычу с самками другой группы, свои самки, взяв кусок, делили его с чужими — и всё без малейших проявлений агрессии! Даже если вдруг окажется, что часть из них была родственницами (что крайне маловероятно, см.выше) уже есть доказательство, что сотрудничество между неродственными самками бонобо «перевешивает» их родство с третьими особями, у этого вида не зависит от родства, а социальная сеть, структурирующая группу, основана на таком дележе и взаимной поддержке, а не на генетической близости особей (из обзора в: Fruth, Hohmann, 2018).
«Политика» у шимпанзе
Вернёмся от ксенофильных бонобо к более или менее ксенофобным шимпанзе. В лесу Тай было показано, что в отличие от восточного подвида, для охраны групповых территорий важны не только самцовые рейды с нападением на чужаков, но и поддержка «держащих» пространство самцов самками группы, выраженная в присутствии, криках, и прочих сигналах urbi et orbi. Собственно, это не новость — во всех притязаниях самцов на более высокий статус, тем более «верховное руководство», поддержка самок более важный момент, чем сила, пыл, характер и прочие качества самих самцов. Поэтому во внутригрупповой конкуренции за лучшее «место» в иерархии самцы тратят куда больше усилий на расположение к себе самок группы и/или снискание их поддержки, чем непосредственно на одоление оппонента.
Именно это де Ваал, изучая долговременную социальную динамику группы шимпанзе в Арнемском зоопарке, назвал «политикой». У всех млекопитающих, где внутригрупповые отношения структурированы системой доминирования-подчинения (обычно агонистического, но бывает и дружественное, основанное на позитивных контактах) именно подчинённые особи проявляют расположение к доминанту, выказывая готовность корректировать свои действия с учётом его активности, в т.ч. прерывать их и отказываться от них (Гольцман, 1983). Не доминант силой устанавливает «право» действовать безотносительно реакции окружающих, но, выказывая подчинение и поддерживая его действия против других особей высокого ранга, подчинённые его к этому «приглашают».
Бонобо всё же не люди: существенные отличия
Не забудем, что бонобо — всё же не человек, в отличие от нас, у него нет «инструментов социального труда», вроде языка и культуры, чтобы «вложить» в каждого понимание необходимости делиться с другими в «кругу равных», или наказывать «обманщиков», действующих в свою пользу, как это «прошито» у всех Homo sapiens; собственно, это стремление к равенству и основанному на нём доверию — «клей», на котором держится всякое общество, а современное капиталистическое — почему не разваливается вопреки классовому антагонизму1 (хотя надо бы, чтобы родилось новое, лучшее, необходимость в котором давно приспела).
Так, в тех же «экономических» играх вроде «Ультиматума» люди, и первобытные и современные, больше заботятся о справедливости дележа, чем о том сколько получат сами. А вот у обоих шимпанзе понятия о справедливости в такой ситуации отсутствуют, они думают о том, как стянуть чужую порцию, а не осталось ли столько же другому (Кaiser et al., 2012). При том что они регулярно делятся пищей, для укрепления социальных связей, и в опытах с обменом пищей разного качества отвергают неэквивалентный обмен (хотя и тут терпимей к неравенству, см. Brosnan et al., 2010)
Так вот, при наличии выбора — делиться с другим или есть в одиночку, — бонобо предпочитали лоханкизм вместо дележа даже со «своими». Это было показано как для хорошо знакомых обезьян в зоопарке Лейпцига, и для малознакомых но из одной клеточной группы в убежище для бонобо-сирото «Lola ya Bonobo».в Конго. Оба вида Pan в отсутствие пищи предпочитали быть в коллективе, но есть — в одиночку. В какой степени обезьяны из одной клеточной группы «чужие» или «свои» — вопрос дискуссионный, но постоянное присутствие по соседству создаёт близость, которая может быть сопоставлена с близостью членов одной группы в природе. Тем не менее надо отметить, что готовность владельца делиться пищей варьировала в зависимости от степени знакомства, при явном предпочтении менее знакомых особей одногрупникам (из обзора в: Fruth, Hohmann, 2018).
Ещё более важная разница продемонстрирована Krupenye et al. (2018). Мы любим тех, кто с нами вежлив, хорош, улыбается; даже трёхмесячные младенцы предпочитают сотрудничающих. А вот бонобо при всём их миролюбии предпочитают тех кто в такой ситуации неудобен, толкается и дерётся. что показывает наши уникальные способности к кооперации.
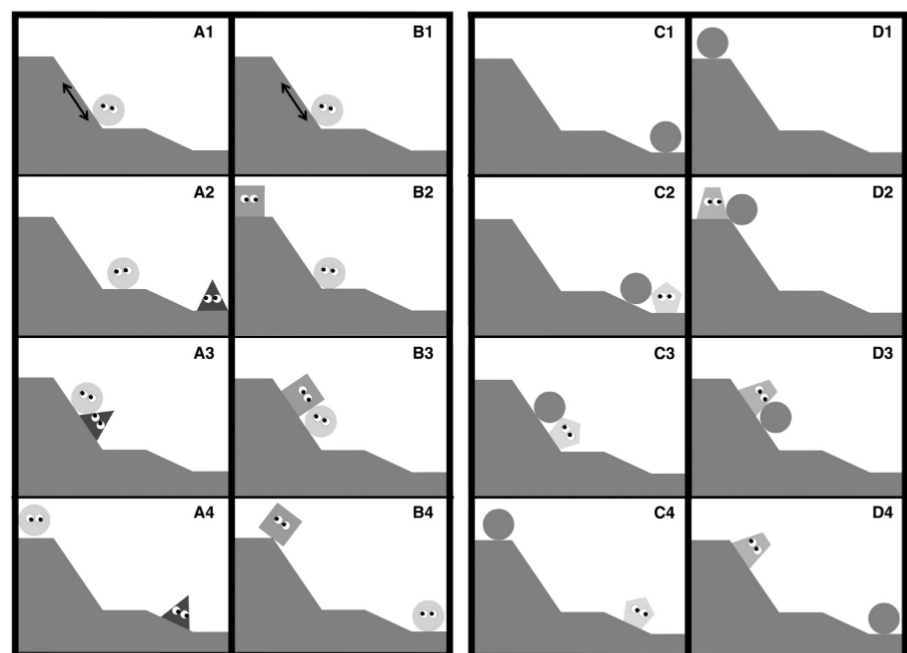
Персонажи из опытов Krupenye et al., 2018 — пробующий забраться на вершину горы, его помощник и антагонист
Что исследовали на 24 бонобо, содержащихся в санктуарии в Республике Конго. Им показывали фильм, чем некий персонаж пробует изо всех сил добраться к вершине крутого холма. Потом показывали картинку, как другой персонаж ему или помогает, или равнодушно смотрит, как он беспомощно карабкается (рис.7). После чего брали лакомство и прятали в конус с одним или с другими персонажем, и смотрели где он будет искать.
Дальше ему показывали скетч, как некто бросает набивную игрушку, другой человек за ними идёт и возвращает а третий, наоборот, выхватывает его и мешает вернуть. После чего обезьяна решала, принять яблоко от «благодетеля» или вора. И во всех этих опытах они выбирали бесполезных или вредных персонажей, причём ещё до того как они начинали себя подло вести. Авторы предположили что они так, потому что грубость — признак высокого статуса, и хорошо иметь таких на своей стороне. Для проверки этой идеи всем 24-м показали мульт, как один персонаж мешает другому занять место. И они таки предпочитали победителя тому кто уступил в борьбе (Krupenye et al., 2018).
Хотя бонобо и «социальнее» шимпанзе, наш уровень просоциальной активности с кооперацией для них недостижим. Здесь нужно согласиться с Майклом Томазелло (цит.по Федорович, 2015), что это результат чисто человеческой инновации — появления производства, труда, и обслуживающих его языка, прочих символических форм культуры. Как давно и прочно показано культурно-исторической теорией Выготского-Лурии-Леонтьева, они «работают» как психические орудия, почему в регулярном применении («практике») прогрессивно развиваются сами и «тянут» за собой наши способности разума и познания, также как выражения того и другого в доступных семиотических системах.
Резюмируем: рассмотрев все относящиеся к делу факты, никак нельзя счесть всю систему характерных особенностей бонобо «самодоместикацией», в том смысле, что создавалась те же селективным давлением, какое одомашнило млекопитающих и было воссоздано в вольерном эксперименте Д.К.Беляева. Отдельные сходства с результатами одомашнивания да, есть, но они созданы явно другими причинами, как показывает рассмотрение всех относящихся к делу фактов.
Городские лисицы и правда самодоместицировались
А вот лисицы Vulpes vulpes, успешно освоившие города Англии и других стран, питающиеся там отбросами, как дворняги, похоже, и правда самодоместицировались (Рarsons et al., 2020, см. пересказ по-русски). Сравнение черепов современных лисиц с только начавшими осваивать город в 1970е показано направленное уменьшение лицевой части черепа, уменьшение среднего объёма мозга и полового диморфизма по черепу.
А при урбанизации, наоборот, мозг «городских» птиц и млекопитающих в сравнении с «сельскими» или лесными скорей увеличивается, как и в других случаях освоения более гетерогенной, нестабильной и изменчивой среды обитания, чем природные ландшафты материка — «островов», созданных фрагментацией этих последних, настоящих океанических островов, и особенно урболандшафтов. Это надёжно показано для птиц, так и для млекопитающих (Sol et al., 2008).
Подчеркну: увеличение мозга вследствие успешной урбанизации не надо путать с идеей, что больший размер мозга вида в сравнении с близкими повышает вероятность успешного освоения городов. При прочих равных нет; вне зависимости от размеров мозга здесь важно изменение жизненного цикла и/или всей жизненной стратегии особей в тех популяциях региона, что испытывают «давление» растущего города, чьи местообитания дробятся, фрагментируются, и нарушаются по краям, и чем дальше тем больше, — от прежнего консерватизма в сторону большей лабильности.
Как только это случается, вид перестаёт «отступать» от такого давления, заселяет всё более фрагментированные и сильней изменённые «острова» своих прежних биотопов, иными словами, «движется» по городскому градиенту «вверх», пока не заселит собственно город. Как и при заселении островов, относительное увеличение мозга может быть следствием последнего, предпосылкой оно оказывается лишь в том случае, когда больший мозг уже сочетается с лабильной жизненной стратегией. Всё это верно для урбанизации не только птиц, но и для млекопитающих (Santini et al., 2019).
Доводы «за» самодоместикацию человека, морфологические и генетические, ещё более разнообразны и значимы. Прежде всего то, что при одомашнивании под отбором оказываются пулы генов, сильно пересекающихся у с генами, бывшими под отбором в процессе антропогенеза (Theofanopoulou et al., 2017; см. рассказ по-русски). Немногое, что не укладывается:
1) в этот период мозг гоминид непрерывно увеличивался, примерно до момента 30000 лет назад, когда мозг Homo sapiens, уже вполне современных, «в среднем» стал уменьшаться;
2) отсутствие в популяциях Homo sapiens подъёма случаев пегости и вислоухости сравнительно с шимпанзе.
Заметки по поводу
Значимость сплочённости внутри группы как целого для победы в конфликте, её несводимость к индивидуальным характеристикам «бойцов», даёт веские доводы, с одной стороны, «против» известной модели коэволюции парохиального альтруизма и войны Боулза и Чо, с другой, «против» идеи группового отбора, точней за достаточность только индивидуального отбора для выработки групповых адаптаций. Когда индивиды не независимы, но связаны между собою в сообщества социальными связями, поддержка одних особей и элиминация других других «сказывается» на целом, отбор в первую очередь меняет формы социальной организации, «используя» перемещения и взаимодействия особей как статистические испытания и расходный материал. Т.е. межгрупповая конкуренция способствует эволюции общего для разных групп социума (разница только в «точности» и и «полноте» реализации его видового «типа» в конкретных условиях существования разных групп) больше, чем изменения качеств самих индивидов. Этот вывод важен и для антропогенеза, не только для обезьян.
Примечание
Свежие комментарии