 Print
Print  PDF
PDF
Резюме. В продолжение темы, как усложнение социальной организации стимулирует прогресс «умственных способностей» у связанных ею особей. На колониальных чистиковых С.П.Харитоновым было показано, что у более социальных видов и исследовательская/манипуляционная активность сложней, птицы проявляют к ней больше интереса. Вовлекаясь в манипуляцию необычными, яркими предметами, птицы начинают исследовать окружающий мир «вообще». См. тупик Fratercula arctica, когда нужно почесаться, целенаправленно берёт острый предмет и чешет грудь. Такое наблюдали и раньше, но на камеру сняли только недавно. См. также описание в PNAS
***
С. П. Харитонов
Резюме авторское. Работу проводили на островах Охотского (о-ве Талан (59°20′ с.ш., 146°05′ в.д.) в 1987–1988, 2008 гг. и о-ве Ионы (56°24′ с.ш., 143°23′ в.д.) в 2009 г.) и Баренцева (о-ве Медвежий, архипелаг Шпицберген (74°23′ с.ш., 19°02′ в.д.), в 2005 и 2011 гг.) морей. У морских птиц отмечен интерес к предметам, не имеющим отношения к поддержанию жизни и размножению. Для выяснения особенностей этого интереса в естественных условиях было проведено 64 опыта с большими конюгами (Aethia cristatella), 47 опытов с люриками (Alle alle), 15 опытов с другими чистиковыми птицами и один опыт с представителем трубконосых – глупышем (Fulmarus glacialis). Птицы обращают повышенное внимание на выделяющиеся из фона предметы, пытаются их исследовать визуальным и тактильным способами.
В наибольшей мере этот интерес отмечен у высокосоциальных морских птиц: люриков и больших конюг. Большие конюги явно отдавали предпочтение предметам белого и желтого цвета. Выраженной также была их реакция на оранжевый и зеленый цвета. Птицы предпочитали более яркие предметы, даже если цвет этих предметов ранее им в природе не встречался. Более яркие предметы стимулировали реакцию конюг на расположенные рядом менее яркие предметы, “сложные” предметы предпочитались “простым”. У люриков, если в опытах был полный набор цветов у одинаковых предметов, реакция на белый цвет заметно преобладала. В опытах, где не было белых предметов, но были одинаковые предметы других цветов, интерес вызывал крайний предмет, какого бы цвета он ни был.
Люрики стремились трогать и перемещать клювом предметы, но уделяли меньше внимания их цвету и форме, чем большие конюги. Сильно пахнущие вещества и вещества с едким вкусом, которые были предварительно нанесены на поверхность предметов, не являлись для больших конюг препятствием к стремлению манипулировать этими предметами. Люрики прекращали попытки манипулировать предметами, если от них исходил запах ментола.
Когнитивное поведение животных изучается преимущественно в лабораториях, где можно поставить довольно строгие опыты с надлежащим соблюдением необходимых условий и обеспечением надежного экспериментального контроля.
В природе опыты по изучению когнитивного поведения проводятся чрезвычайно редко (например, Обозова и др., 2010). Непосредственно в тех местах, где животные обитают, экспериментаторы встречаются с рядом трудностей. Прежде всего, в природе, в отличие от лаборатории, невозможно обеспечить строгий контроль. Не всегда можно обеспечить и постоянство условий (может меняться погода; могут сменяться особи, участвующие в экспериментах; не всегда возможно определить физиологическое состояние особей и пр.).
Главная трудность состоит в том, что условия проведения опытов в природе приходится приводить в соответствие с условиями опытов, проведенных в лаборатории, пусть и с другими видами. Выход здесь представляется такой: где возможно, все же стараться соблюсти постоянство условий; при малейшей возможности пытаться осуществить контроль в опытах; постараться вынести из результатов как можно больше аргументов в пользу предлагаемой интерпретации.
Есть еще одна важная особенность лабораторных экспериментов: для того чтобы животные выполнили требуемое задание, их мотивируют возможным вознаграждением (де Вааль, 2017). В природе создать мотивацию вознаграждением обычно не удается (хотя иногда такое получается – Обозова и др., 2010), поэтому остается прослеживать, как животные выполняют задание сами по себе, не ожидая вознаграждения от человека. Нам представляется, что принцип “добровольности” – это очень важная особенность полевых экспериментов, которая подчас помогает лучше понять не только умственные способности животных, но и те направления деятельности, к которым они свои умственные способности прикладывают вне круга привычных жизненных адаптаций.
Как известно, важнейшую форму когнитивного (познавательного) поведения животных составляет исследовательская активность, которая обеспечивает знакомство особи с окружающей средой, с происходящими в ней изменениями и со свойствами окружающих и особенно новых предметов. Этологи (Дьюсбери, 1981; Обозова и др., 2013) рассматривают исследовательскую активность как необходимый компонент в репертуаре индивидуального поведения (наряду с кормовым, локомоторным, терморегуляционным, оборонительным и др.). Широко распространенным способом ознакомления со свойствами предметов является манипулирование ими, в связи с чем этот вид когнитивной деятельности оценивают по проявлениям манипуляционно-исследовательской активности.
В качестве объекта исследований были выбраны морские птицы, в основном, два их представителя: большие конюги и люрики. Эти птицы гнездятся в крупноблочных осыпях, в пустотах под камнями и обладают очень сложной социальной системой, отличной от таковой у других видов чистиковых птиц (Харитонов, 2006; 2011). На о-ве Талан в 1987–1988 гг. мы наблюдали, как большие конюги обрывают лепестки лапчатки, а также реагируют на необычные для них предметы на колонии: ощупывают клювами некоторые участки камней, хватают клювами цветные кольца друг на друге, ощупывают клювом блестящие металлические предметы и т.п. Лепестки цветов и мелкие пластиковые предметы найдены нами в гнездовых камерах больших конюг. Подобное поведение наводило исследователей на мысль разместить на поверхности осыпи, где гнездятся большие конюги, небольшие цветные предметы и посмотреть на реакцию этих птиц. Первые такие исследования были проведены в 1990 г. В.А. Зубакиным и Е.В. Зубакиной (Зубакин, Зубакина, 1990) с набором небольших цветных дисков оранжевого, красного, желтого, зеленого, светло- и темно-коричневого и белого цветов. Птицы манипулировали клювом дисками всех цветов, однако первая реакция больших конюг показала наличие у этих птиц неких цветовых предпочтений: первая реакция отмечена на белый и оранжевые цвета. На первый взгляд, такая реакции представляется вполне объяснимой. Оба эти цвета имеют для больших конюг сигнальное значение в брачный сезон: у них ярко-оранжевый клюв, контрастная белая косица на черной голове и белая радужина глаза (Зубакин, Зубакина, 1990).
Люрики на о-ве Медвежий тоже рассматривали и ощупывали клювом поверхность камней, трогали и даже тянулись клювом к деталям ловушек. Подобное поведение побудило нас провести опыты по выяснению реакции люриков на цветные предметы, разложенные на камнях в колонии. Наблюдения и опыты позволили собрать материал об исследовательской активности и других чистиковых птиц, гнездящихся рядом с большими конюгами. Малые конюги (Aethia pygmaea), конюги-крошки (A. pusilla), белобрюшки (Cyclorrhynchus psittacula), ипатки (Fratercula corniculata) и топорки (Lunda cirrhata) тоже образуют клубы на камнях, однако уровень социальных взаимодействий у этих видов слабее (Харитонов, 2006; 2011). Белобрюшки изредка хватали клювом пятна накипных лишайников, ипатки и топорики – помет на камнях, а ипатки – также растущие зеленые травинки.
Многие птицы способны чувствовать запах (Поповкина и др., 2010). Подобная чувствительность обнаружена и у больших конюг (Hagelin, 2007; Hagelin et al., 2003). В данном исследовании мы попытались, используя интерес больших конюг и люриков к выложенным на камнях колонии предметам, определить реакцию птиц на некоторые запахи, которые с точки зрения человека резки и неприятны.
Материалы и методы
Содержание
Работу проводили на островах Охотского моря: о-ве Талан (59°20′ с.ш., 146°05′ в.д.), в 1987–1988, 2008 гг. и о-ве Ионы (56°24′ с.ш., 143°23′ в.д.) в 2009 г.; и Баренцева моря: о-ве Медвежий, архипелаг Шпицберген (74°23′ с.ш., 19°02′ в.д.), в 2005 и 2011 гг. На камни, где проходила клубная активность птиц, выкладывали небольшие предметы: разноцветные пластиковые кольца для мечения птиц, большие и малые канцелярские скрепки с цветной оплеткой, подсвечники от мелких свечек, женские заколки, кусочки цветного пластика, яркие овальные предметы (продающиеся в магазинах небольшие вешалки для одежды), пластиковые крышки от бутылок для воды, яркие пластиковые диски диаметром до 15 см.
Предметы предъявлялись птицам обычно на периоды от 10 мин до 1 ч, затем убирались одни предметы и могли быть предъявлены другие. С большими конюгами в 2008 г. на о-ве Талан проведено 64 опыта (предъявлений) со всеми вышеперечисленными предметами. С люриками на о-ве Медвежий в 2005 г. проведено 5 экспериментов. В этих опытах использовались только цветные кольца для птиц. В 2011 г. с люриками проведено 42 опыта, в которых использовались разнообразные перечисленные выше предметы. На о-ве Ионы в 2009 г., кроме выкладывания разноцветных скрепок и цветных колец, использовался небольшой красный диск диаметром 5 см, подвешенный на веревке возле самой поверхности камня. На о-вах Межвежьем и Ионы некоторые предметы в опытах приклеивались к камням суперклеем, и они фактически постоянно находились на поверхности колонии. Во время наблюдений записывали реакцию птиц на эти предметы.
Семь опытов проведено с ипатками, четыре опыта – с топориками, 4 опыта – с малыми конюгами и конюгами-крошками. Еще один опыт, где использован большой пластиковый красный диск диаметром 15 см – с глупышем. Если птица целенаправленно касалась предмета, то считалась, что предмет привлек ее внимание, и реакция на него положительная. Чаще всего, однако, касанием дело не ограничивалось – птицы начинали при помощи клюва манипулировать предметами. Ход всех опытов протоколировался видеосъемкой камерами JVC‐270E и JVC‐340Е, обладающими большим оптическим увеличением, что позволяло при необходимости настолько приблизить изображение, чтобы были видны даже малозаметные детали процесса манипулирования предметами. Всего отснято более 22 ч наблюдений и опытов с большими конюгами и около 18 ч – с люриками. Протоколы опытов составлялись на основе видео и записей в дневнике, поскольку не все реакции удалось отснять, некоторые удавалось отснять частично – наводили камеру тогда, когда птица уже трогала предмет. В случаях, когда сам момент прикосновения не удавалось записать на видео, сведения брались из дневника. Если птица хотя бы касалась предмета клювом, а тем более – манипулировала им, то это считалось положительной реакцией на данный предмет. При сравнении реакции на разные предметы подсчитывалось число касаний и манипуляций. Отдельно рассматривались первые касания в каждом опыте и общее число касаний в течение опыта. Хотя число проведенных опытов измеряется всего лишь десятками и, соответственно, десятками могло быть измерено число первых касаний, общее число птиц, коснувшихся предметов, и общее число касаний исчислялось сотнями.
При обработке результатов опытов предметы разносились по категориям. В опытах, где изучалась реакция птиц на разные цвета, одной категорией считались предметы одного цвета. В таких опытах использовались одинаковые по форме предметы разного цвета (например, только цветные кольца или цветные канцелярские скрепки). Хотя в каждом отдельном опыте предметы были одинаковые, при выяснении реакции на разные цвета опыты с предметами разной формы могли быть объединены в одну выборку. В опытах, где изучалась реакция на форму и яркость предметов, использовались предметы разной формы; были выделены категории: “простой предмет”, “сложный предмет”, “яркий предмет”, “неяркий предмет”.
Один и тот же предмет в разных опытах мог быть отнесен к разным категориям, в зависимости от того, какие еще предметы одновременно предъявлялись. Например, если в опытах был выбор между цветными кольцами для птиц и подсвечниками для елочных свечей – кольца считались “простым” предметом, а подсвечники – “сложным”. Если был выбор между теми же подсвечниками и фигурными женскими заколками для волос, то здесь уже подсвечники считались “простым” предметом, а заколки – “сложным” предметом (рис. 1).
В 16 экспериментах с большими конюгами на поверхность предметов наносились различные химические агенты с резким запахом. В качестве запаховых агентов использовались: ментол, содержащийся в средстве от комаров “Москитол” и “Тигровой мази”, 10% раствор аммиака, измельченный зубчик чеснока, вазелин с протертым чесноком и выжатый лимонный сок. Последний взят потому, что вещество, которое выделяется большими конюгами и которое (как считается) обуславливает этот запах, содержит компонент с запахом цитрусовых (Hagelin, 2007). С люриками в 2011 г. в 7 опытах использовали запаховые агенты двух типов: издающие запах аммиака – 10% раствор этого вещества и издающие запах ментола – ментоловое масло, средство от комаров “Москитол” и “Тигровая мазь”.
У больших конюг по размеру и форме клюва довольно легко можно визуально определить пол птиц (Конюхов, 1996; Jones et al., 2004), поэтому в большинстве случаев удавалось понять реакцию на предметы птиц разного пола. Люрики значительно более мономорфные птицы, чем большие конюги, у них определить пол в полевых условиях чрезвычайно трудно. В последние годы появилась весьма надежная методика определения пола птиц по фотографиям их клювов в профиль (Виноградов, 2015). Во время нашей работы эта методика отсутствовала, хотя в ряде случаев она нам позволила определить пол люриков по видеозаписям. Однако в те годы съемка велась еще камерой стандартного разрешения (720 × 576 пикселей), что во многих случаях было недостаточно для получения полноценных стоп-кадров и точных измерений пропорций клюва. Поэтому при обработке результатов пол люриков, в основном, не учитывался.

Рис. 1. Предметы, использованные в опытах: a – цветные кольца для птиц; b – канцелярские скрепки с цветной оплеткой; c – кольца и подсвечники; d – заколки, подсвечники и малый красный диск; e – кольца и заколки; f – заколки и подсвечники; g – кольца и заколки; h – заколки и подсвечники; i – разноцветные вешалки.
Для статистического анализа использовали тест для альтернативных признаков с поправкой на неоднородность выборок (Лакин, 1990), а также сравнение выборочных долей с генеральной средней (Плохинский, 1978). В опытах на цветовые предпочтения рассматривалось 6 цветов, поэтому выбор цвета считался случайным, если число реакций на него недостоверно отличалось от 1/6 (генеральная средняя в этом случае равна 0.1666667). Если в опытах на предпочтение цветов было не 6, а только 3 цвета, то, соответственно, генеральная доля для случайных касаний составляла 0.3333. В случае выбора из двух альтернатив (яркий–неяркий предмет, “простой”–“сложный” предмет) случайным выбором считалось недостоверное отличие от 1⁄2 (генеральная доля 0.5).
Результаты
Большие конюги и люрики толкали предметы клювом, брали их в клюв, могли положить под себя на плавательную перепонку или бросить через плечо. Видимо, некоторые из этих движений являются элементами гнездостроительного поведения, хотя конюги никакой подстилки в гнездо не приносят.

Рис. 2. Фото большой конюги, касающейся предметов.
Люрики, однако, приносят подстилку в гнездо в виде небольших плоских камней (Stempniewicz, 1986; Cramp, 1985). Обычно птицы просто перетаскивали предметы с места на место (рис. 2). Вся эта деятельность с предъявляемыми предметами вполне подходит под понятие “манипуляционная деятельность”, которая наблюдается у млекопитающих (Дерягина, 1986). Однако в данном случае нам кажется более подходящим термин – манипуляционно-исследовательская активность (Зорина и др., 1986). Поскольку экспериментатор ничем не побуждает птиц манипулировать предметами, эту активность птиц вполне можно считать спонтанной.
Реакция больших конюг на предметы.
Наблюдая за тем, какие предметы конюги трогали клювом, брали в клюв и манипулировали ими, мы сделали попытку проанализировать закономерности этой манипуляционной активности. Одна из задач опытов – выяснить цветовые предпочтения конюг вне зависимости от категории предметов. Полный экспериментальный набор содержал предметы следующего цвета: белый, желтый, оранжевый, красный (разные оттенки, от розового до ярко-красного и темно-красного, т.е. бордового), голубой (разные оттенки), зеленый (разные оттенки), черный. Поскольку наибольшее число опытов проведено с предметами, окрашенными в 6 цветов (таблица, рис. 3), по этим цветам и проведен основной анализ.
Полный набор цветов использовался в 19 опытах, где предметы были небольшими и неяркими. Первая реакция зафиксирована 16 раз у самцов и 14 раз у самок. Всего же у самцов отмечено 112 случаев касаний и манипулирования предметами, у самок – 47. Белый, желтый и оранжевый цвета были для конюг более предпочтительны, чем зеленый, синий и красный (рис. 3). Первая реакция самцов на желтые предметы достоверно превышала уровень случайной реакции на этот цвет (таблица). Если же рассматривать все реакции, то у самцов достоверно преобладала реакция на предметы белого и желтого цветов (таблица). У самок первая реакция на белый и желтый тоже преобладала, однако, на более низком уровне достоверности (таблица). В случае, если предъявлялись небольшие предметы многих цветов, то уровень всех реакций самцов на предметы красного цвета был значительно и достоверно слабее, чем в ситуации, если бы касание предметов было случайным (таблица).
У самок по общему числу реакций получилось преобладание белого цвета, и, в то же время, более отчетливая, чем у самцов, первая реакция была на оранжевый цвет (таблица, рис. 3), который, скорее всего, имеет для этих птиц сигнальное значение, поскольку рамфотека больших конюг оранжевого цвета.
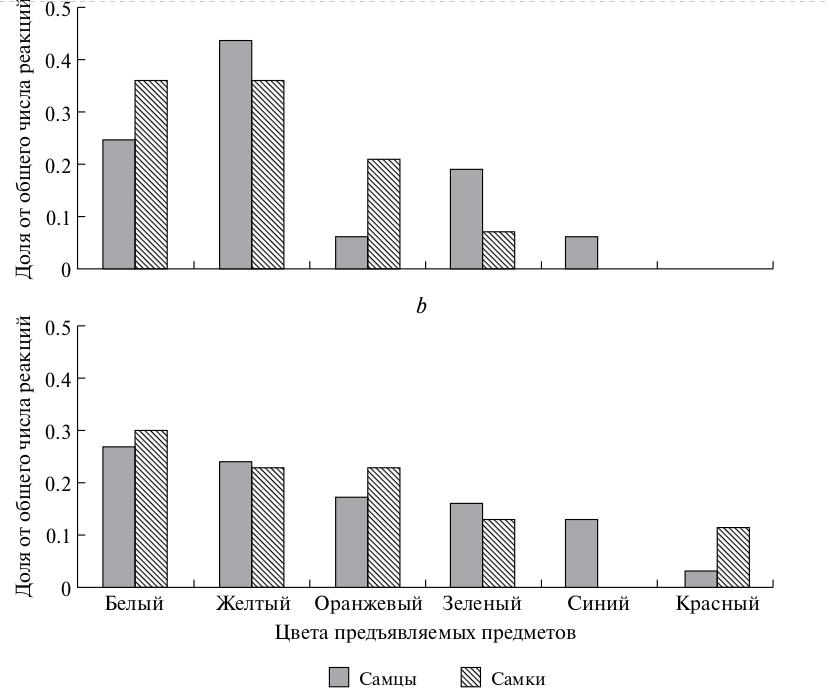
Рис. 3. Распределение цветовых предпочтений у больших конюг: a – первая реакция на предметы, b – все реакции на предметы.
Для более детальной обработки мы рассмотрели по отдельности три наиболее предпочитаемых цвета (белый, желтый и оранжевый) и три менее предпочитаемых цвета (зеленый, синий и красный). Это дало возможность увеличить выборку, включив в нее опыты с теми же предметами, но с неполным набором цветов. В результате число опытов, где рассматривалась реакция на три предпочитаемых цвета, достигло 29, на три менее предпочитаемых цвета – 24. Количество опытов и реакций на них позволило отдельно рассмотреть два временных периода – реакции птиц до 1 июля и после.
Разделение на такие временные периоды связано с тем, что с самого начала июля у больших конюг начинается линька рамфотеки, и клювы резко теряют яркость оранжевого цвета. До 30 июня включительно у самцов при выборе из 3 предпочитаемых цветов самое сильное и достоверное предпочтение проявилось при первой реакции на желтый цвет (таблица). После первого июля и у самцов, и у самок резко и достоверно усилилась реакция на оранжевый цвет, по сравнению с реакцией на белый и желтый, которая у обоих полов практически не изменилась. Первая реакция самцов на оранжевый в период до 1 июля была почти нулевая, тогда как с 1 июля самцы реагировали на оранжевый цвет почти в половине всех первых реакций (Р = 0.012). Если рассматривать все реакции самцов, то усиления их предпочтения оранжевого цвета не обнаруживается. У самок, как в первой реакции (0 из 10 до 1 июля и 7 из 10 после 1 июля, P = 0.002), так и во всех реакциях (2 из 26 до 1 июля и 15 из 31 после 1 июля P = 0.0015), достоверно возрастает интенсивность реагирования на оранжевый цвет по ходу сезона.
При этом у самок резко уменьшилась доля реакций на желтый цвет: 13 из 26 всех реакций до 1 июля и 7 из 31 всех реакций после 1 июля (таблица). Среди трех реже выбираемых цветов (зеленый, синий, красный) у самцов проявилась резко негативная реакция на красный цвет. Здесь мы сравнивали только все реакции птиц, поскольку результаты первых реакций составили малые выборки. До 1 июля у самцов было 4 из 38 против ожидаемого 12.5; после 1 июля характер реакции не изменился – 1 реакция из 7 зафиксированных (таблица).
У самок реакция на небольшие красные предметы была значительно сильнее, чем у самцов, близка к случайному уровню, после 1 июля интенсивность возросла до 0.5 от всех реакций, но недостоверно. В 13 опытах рядом с ранее применяемыми предметами (цветные кольца или скрепки) подкладывались ярко-красные диски разного размера.
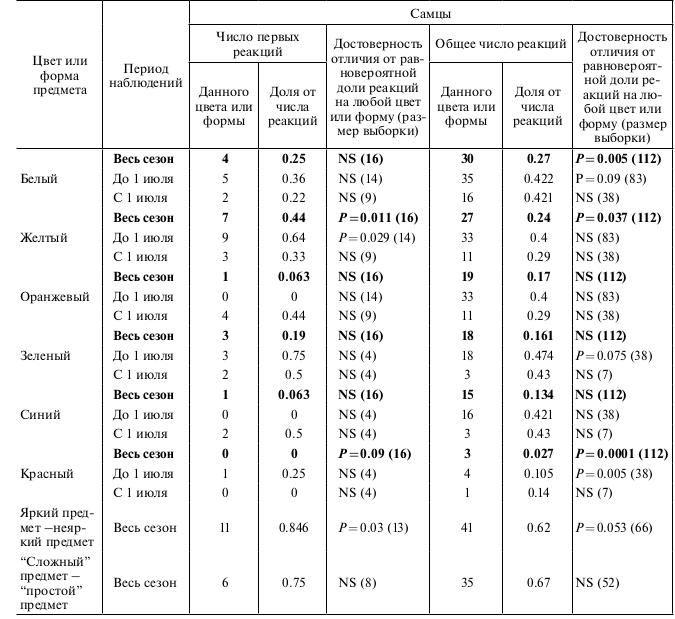
Таблица. Число реакций на предметы разного цвета и формы у больших конюг. Примечания. Жирным шрифтом выделены результаты опытов, где предъявлялось 6 цветов, здесь доля случайных реакций считалась 1/6 (0.1666667) от общего числа реакций. В результатах опытов по выбору цветов, которые представлены обычным шрифтом, конюгам предъявлялось или рассматривалось по 3 цвета в каждом опыте, Случайной долей здесь считалась 1/3 (0.3333333). В опытах на яркость и форму предметов учитывался выбор из двух альтернатив, поэтому случайным считался выбор, если он недостоверно отличался от 1⁄2 (0.5).
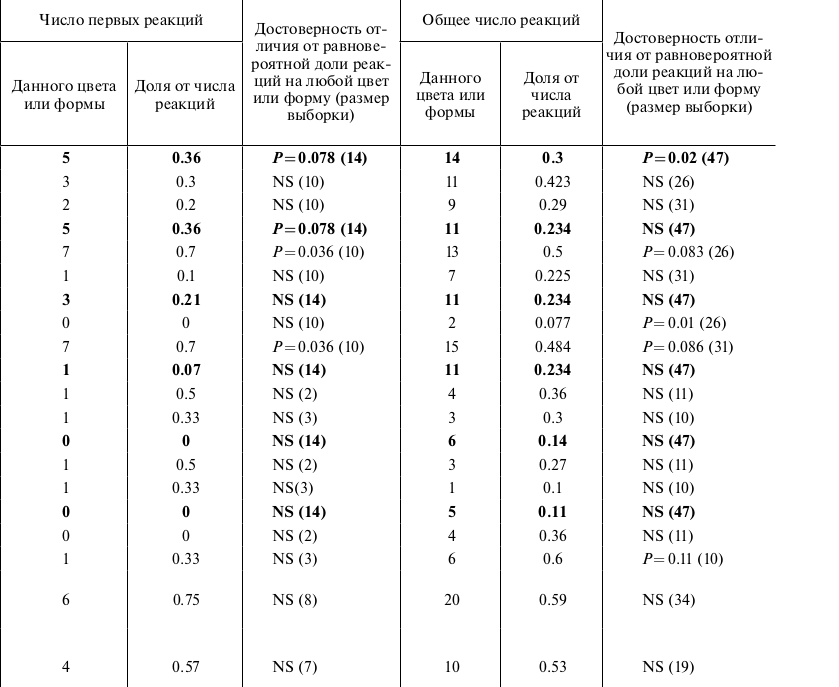
Оказалось, что наличие яркого крупного предмета коренным образом изменяет реакцию и самцов, и самок больших конюг. Несмотря на то, что диски были наименее предпочитаемого красного цвета, птицы их достоверно чаще выбирали и при первой, и в последующих реакциях (таблица). При объединении числа реакций у птиц обоих полов данная тенденция прослеживается значительно сильнее: первая реакция 17 из 21 реакций были на яркий предмет, P = 0.01; для всех реакций – 61 из 100, P = 0.03. У самцов эта реакция на яркий большой предмет была выражена значительно сильнее, чем у самок (таблица). Т.е. яркие крупные предметы не встречающегося в природе цвета оказались для больших конюг предпочтительней, чем предметы знакомых цветов. Еще в 8 опытах были добавлены предметы черного цвета.
На черный цвет птицы реагировали хуже всего и только тогда, когда рядом были предметы других цветов. Реакция на черный цвет была явно сходна с выбором: “яркий предмет – неяркий предмет”, где черный играл роль неяркого предмета. На отдельный черный предмет, без других цветных предметов рядом, конюги не реагировали. Это означает, что более яркие предметы еще и привлекают внимание к менее ярким. Если конюгам предлагалось несколько ярких красных предметов (диски и пластиковая полоска), они выбирали не более крупный предмет, а тот, который оказывался к ним ближе (3 опыта): 25 реакций на ближе расположенный предмет и полное отсутствие реакций на немного более удаленный предмет, хотя он тоже был большой и яркий.
В 9 опытах большим конюгам предлагался выбор из категорий: менее “сложный” предмет – более “сложный” предмет. “Сложность” и “простота” определялась следующим образом: птицам предъявляли предметы примерно одного размера, но одни с более сложным изрезанным контуром поверхности, чем другие. Результаты этих опытов были не столь выразительны, как в случаях с яркими крупными предметами. Однако у конюг обоего пола прослеживалась четкая тенденция к предпочтению более “сложного” предмета (таблица). В первой реакции разница недостоверна даже при объединении данных от обоих полов; среди всех реакций при объединении данных от птиц обоего пола (71 реакция), в 45 случаях птицы предпочитали более сложный предмет, P = 0.027).
При выборе между “ярким предметом” небольшого размера (малый красный диск) и “сложным” предметом сходного с ярким предметом размера (цветные женские заколки для волос) не обнаружилось предпочтений одной из этих категорий (11 реакций на яркий предмет, 12 – на “сложный”), хотя здесь выборки малы для полноценного анализа. В целом, из небольших относительно простых предметов (скрепки или кольца) первым берут предмет определенного цвета (результаты выше), а из ярких предметов – любой, который ближе, вне зависимости от его размера. Замечены также следующие особенности: при наличии плоских и объемных предметов птицы обычно трогают объемные предметы, например куски торфа над оранжевой полоской и черный камень над оранжевой полоской.
Реакция люриков на предметы.
Реакцию люриков на предметы оказалось гораздо труднее систематизировать, чем реакцию конюг. Здесь относительно выше была доля опытов, где люрики ни разу не коснулись предметов (у конюг – 3 опыта из 64, у люриков 9 из 47, хотя если из выборки убрать опыты с запаховыми агентами (см. ниже), где некоторые запахи вызывали заметное для наблюдателя отвращение люриков, то люрики не касались предметов в 6 опытах из 40). При этом движения намерения коснуться отмечались у них почти в каждом опыте. У люриков нет такой быстроты в хватании предметов, как у конюг, они более осторожны и обычно дольше “примериваются”. Реакция по сравнению с конюгами – замедленная, нередко представлена “порциями”: сначала увидел предмет, посидел. И только потом начал действовать дальше – тянуться и трогать. Несколько раз утаскивали предмет под камни.
Люрики, так же как и большие конюги, манипулировали клювом с предметами всех цветов, но первая реакция была четкой: если в опытах был полный набор цветов у одинаковых предметов, реакция на белый цвет явно преобладала (11 опытов за два сезона, первая реакция на белый 8 против 2 – на бордовый, всего – 22 реакции на белый из 53, отличие от случайного P = 0.00001). В опытах, где к небольшим одинаковым предметам (кольца для птиц или цветные скрепки) подкладывали красные яркие пластиковые диски, из пяти таких опытов в трех реакция была на красные диски, в двух опытах – на белые мелкие предметы. В опытах, где не было белых предметов, но были одинаковые предметы разных цветов, птицы отчетливо касались сначала крайнего предмета и манипулировали им, какого бы цвета он ни был: первая реакция в разных опытах была на черный, синий, красный, последующие реакции были на бирюзовый, зеленый, синий, черный и красный.
Люрики интенсивно трогали и перемещали клювом предметы, но гораздо меньше, по сравнению с большими конюгами, обращали внимание на различия предметов по цвету и форме. Обычно они сначала брали крайний предмет, не важно, какой он был, а уж потом хватали из середины области, где были разложены предметы. Возможно, это было связано с относительно меньшей подвижностью люриков по сравнению с большими конюгами. Наши данные не выявили предпочтений по категориям: “простой” предмет – “сложный” предмет; “большой яркий” предмет – “простой неяркий” предмет и другими.
Опыты с другими видами морских птиц.
Начать опыты с другими видами морских птиц нас побудила реакция некоторых ипаток во время опытов с большими конюгами. В одном из опытов с конюгами одна из ипаток, которые совершенно не интересовались предметами, увидев суету конюг возле предметов, вдруг приблизилась к разложенным на камне цветным скрепкам и, наклонив голову, стала их рассматривать. На следующий день одна из ипаток во время пятого опыта с большими конюгами вдруг потянулась к цветным скрепкам явно с намерением дотронуться, но так и не дотронулась. В предыдущих четырех опытах, проведенных с большими конюгами в этот день, ипатки на предметы не реагировали.
В опыте 28.06.2008 ипатка дважды коснулась подложенного нами черного камня диаметром примерно 5 см. Этим камнем мы для опытов с конюгами придавили полоску из оранжевого пластика длиной примерно 12 см. В этот день численность ипаток на камнях была высокой, и конюги, которые значительно меньше ипаток по размеру, боялись подойти к выложенным предметам. 29.06.2008 самка ипатки взяла желтую скрепку и перенесла ее, затем она еще раз коснулась этой скрепки, посмотрела вниз с камня, на колышущуюся на ветру зеленую траву. Далее она пошла по камню к куртине травы, растущей рядом, и стала теребить клювом травинки (до этого зеленую траву не теребила). В опыте 09.07.2008 самец ипатки дважды взял бордовую, затем светло-зеленую скрепку, уронил ее, затем пошел в конец камня (куда и самка 29.06.2008) и стал теребить травинки, растущие возле него. До хватания скрепок травинки клювом не теребил. В этом же опыте самка ипатки несколько раз трогала клювом бордовый пластиковый ромб длиной 7 см, лежавший в лужице лимонного сока (один из опытов на запаховую чувствительность больших конюг).
15.07.2008 ипатка несколько раз трогала этот же бордовый ромб, положенный на камень в опытах с топориками, в другом опыте ипатка несколько раз трогала большой (15 см в диаметре) красный диск. А 14.07.2008 ипатка устроила настоящее “цирковое” представление со средним (8 см в диаметре) и большим красными дисками. Здесь ипатка, кроме всего прочего, дважды трогала себя за ногу.
Топорики оказались наиболее пугливыми по отношению к ярким предметам. К большому красному пластиковому диску они откровенно боялись подходить (два опыта). Когда им предъявили небольшой (до 7 см длиной) не столь яркий бордовый пластиковый ромб, они его несколько раз касались клювом (два опыта). Это максимальная реакция на предметы, которую удалось наблюдать у топориков.
На о-ве Ионы 08.07.2009 к наклонной поверхности камня (по таким обычно перемещаются малые конюги) были приклеены одна большая и 4 малые разноцветные канцелярские скрепки и 2 небольшие фигурные заколки для волос. 09.07.2009 утром малые конюги и конюги-крошки не реагировали на приклеенные предметы, даже если сидели рядом с ними. Большие конюги – наоборот, специально спускались с вершины камня, чтобы потрогать предметы и даже хватали их клювом из неудобного для себя положения, пытались оторвать от камня. Малые конюги в этот день не реагировали, но теоретически могли бы, поскольку одна малая конюга интересовалась пятном помета на камне. Подобного интереса к естественным пятнам на камнях у конюг-крошек ни разу не отмечалось. Вечером 09.07.2009 на тот же наклонный камень при помощи белой пластиковой веревки прикрепили малый красный диск диаметром 5 см.
Большие конюги сразу же стали его трогать; взрослый токующий самец в перерывах между хватаниями диска исполнял основную токовую песню одиночного самца (Харитонов, 1980). Конюга-крошка, сидевшая рядом, не реагировала на диск. Только с утра 10.07.2009 и малые конюги, и крошки (последние, может быть, даже в большей степени) вдруг заинтересовались малым красным диском и белой пластиковой веревкой и стали трогать их.
Причем и у крошек, и у малых наблюдается эффект “наведения”: видя перед собой красный диск, они начинают трогать камни; крошки в обычной ситуации камни клювом не ощупывали. Одна из малых конюг в этой ситуации даже тронула приклеенную к камню бордовую скрепку.
Отмечено, что даже глупыши ощупывают клювом камни, но только по пути своего следования по колонии, направленного движения глупышей к предметам не отмечали. Они трогали даже такие новые предметы, как части ржавой генераторной установки АБ‐4. Они ее ощупывали по пути своего следования, действительно проявляя видимый интерес. В опыте теребили клювом большой красный диск диаметром 15 см, специально положенный на камень. Однако обследовали предметы буквально несколько секунд, не более. Такого упорства, какое характерно для больших конюг, здесь не отмечено.
О запаховой и вкусовой чувствительности у чистиковых птиц.
Поскольку у больших конюг отмечена чувствительность к запахам (Hagelin et al., 2003), в ряде своих опытов мы использовали и запаховые агенты. В двух опытах 6 цветных колец, предъявленных большим конюгам, были политы москитолом. В одном опыте на красный пластиковый диск диаметром около 8 см была нанесена тигровая мазь, в другом опыте тигровая мазь была нанесена на цветные кольца, еще в одном опыте – на большие цветные канцелярские скрепки. В двух опытах также использовали красный диск и кольца для птиц, однако, кроме тигровой мази, предметы обильно полили 10% раствором аммиака. В 6 опытах поверхность пластиковых канцелярских скрепок натерли измельченным чесноком, причем в 4 из них – натертый чеснок смешивали с вазелином для придания стойкости запаху. В трех опытах предметы поливали лимонным соком, в двух случаях из упомянутых трех – густой лимонный сок смешивали с вазелином.
Чтобы точнее описать реакцию конюг на предметы, обработанные запаховыми агентами, необходимо упомянуть характерную для этих птиц поведенческую особенность: эти птицы временами встряхивают клювом, очевидно сбрасывая с надклювья капли воды с повышенным содержанием соли, которые выделяют солевые железы этих птиц.
Реакция на вещества, издающие резкий запах, у больших конюг была такова. Не отмечено никакой специальной реакции на агенты, имеющие запах ментола (москитол и тигровая мазь) и запах лимона (густой свежий лимонный сок). Большие конюги продолжали хватать такие предметы примерно с той же интенсивностью, как предметы вообще без запаховых агентов. Увеличения частоты встряхивания клювом также визуально отмечено не было. Реакция на сильный запах аммиака отмечена, но очень слабая. Птицы все равно хватали предмет, облитый нашатырем или смесью нашатыря с тигровой мазью, но хватали его, иногда сначала отдергивая клюв, или значительно чаще, чем обычно, также встряхивали клювом после хватания диска. При этом они не прекращали хватать предмет в дальнейшем, даже после того, как, судя по такому поведению, явно почувствовали запах аммиака.
В опытах с чесночной пастой птицы никак не проявили запаховой чувствительности к чесноку, зато, как нам представляется, отчетливо проявили чувствительность вкусовую. Птицы, которые не хватали предметы, натертые чесночной мазью, а лишь касались их, не демонстрировали никакого отличия в поведении от тех опытов, где предметы не были ничем натерты. Птицы, которые хватали предмет раскрытым клювом, затем демонстрировали резкое неоднократное мотание головой из стороны в сторону значительно чаще и сильнее, чем это птицы делают, избавляясь от выделений соли из солевых желез. Хотя опытов проведено мало, отмечено отличие полов в восприятии вкуса чеснока: самки ни разу не мотали головой после взятия в рот предметов, натертых чесночной пастой (3 случая взятия в рот самками натертых чесноком предметов), самцы, однако, в 4 случаях из 5 долго встряхивали головой.
В опытах с люриками раствор аммиака использовали лишь в одном опыте, когда птицам были предъявлены облитый нашатырем красный диск диаметром примерно 5 см (тот же диск в подобном опыте с конюгами на о-ве Талан) и пропитанная нашатырем бумага. Люрики во время опыта ходили по этим предметам, не проявляя никакой реакции на запах аммиака. В остальных 6 опытах использовались агенты, издающие сильный запах ментола.
При этом в трех из этих опытов все предъявляемые предметы были облиты веществами с запахом ментола, в трех других опытах часть поверхности предметов была обработана веществами с запахом ментола, а часть нет. Реакция на ментол у люриков была вполне однозначная: они могли тянуться к предметам, пахнущим ментолом, но ни в одном случае не брали их клювом. Нередко наблюдалось резкое отдергивание клюва от такого предмета. Предметы без ментолового запаха люрики хватали клювом так же, как и в других опытах, где не использовались запаховые агенты.
Обсуждение
Реакция больших конюг и люриков на цвет и форму предметов.
Интерес к цветным мелким предметам и манипулирование ими при помощи клюва известно у попугаев, в частности, у волнистых попугайчиков в неволе (Pepperberg, 2008 по: Вааль, Франс де, 2017), а также у молодых врановых (Зорина и др., 1986). Опыты в естественной среде показали, что морские птицы разных видов, из которых рассмотрено 6 видов чистиковых и один вид трубконосых, проявляют интерес и при помощи клюва манипулируют предметами, не имеющими отношения ни к пищевым объектам, ни к процессу размножения. Особенно отчетливо и разнообразно эта реакция проявилась у двух видов: больших конюг и люриков. Для этих двух видов характерна высокая социальная активность во время всего сезона размножения и сложная социальная структура колоний (Харитонов, 2011).
Реакция птиц на предъявляемые предметы говорит об их интересе к окружающему миру, интересе, явно выходящем за пределы повседневных необходимых (адаптивных) действий. Птицы здесь явно проявляли манипуляционно-исследовательскую активность, некоторое стремление познавать окружающий мир. Что-то необычное вызывало у них активный интерес и стремление это “узнать”.
Большие конюги явно отдавали предпочтение предметам белого и желтого цвета, как в первых, так и в последующих реакциях. Значительное внимание уделялось также оранжевым и зеленым предметам. Большие конюги предпочитали яркие предметы “простым” и менее ярким, даже если те были не встречающегося в природе цвета.
“Сложные” предметы предпочитались “простым”. Яркие предметы стимулировали реакцию на рядом расположенные менее яркие. Так, трогали черный камень, если рядом были более яркие предметы, но не трогали его, если рядом лежал обычный (серый) камень, серый камень тоже не трогали. Точно так же и у ипаток – наличие новых предметов стимулировало манипуляционную активность вообще: птицы начинали трогать клювом камни, а также хватать клювом зеленую траву и даже собственную цевку.
Можно отметить также проявление такого феномена как “социальное облегчение” (обзор Резникова, 2005), когда манипуляционная активность конюг направляла внимание ипаток на предметы и стимулировала манипулирование ими. Люрики и конюги своей реакцией на предметы стимулировали также других особей своего вида, которые сами до этого не интересовались предметами.
В наших опытах, в отличие от опытов, проведенных в 1989 г. (Зубакин, Зубакина, 1990), проявилась сильная реакция больших конюг на предметы желтого цвета, чего не выявлено в упомянутых более ранних опытах, хотя предметы желтого цвета в изучаемом наборе там были. Судя по реакциям птиц, в наших опытах желтый цвет активно воспринимается практически наравне с белым.
Объяснить подобное различие можно следующим образом. Конюги нередко трогают засохшие пятна помета на камнях. Помет больших конюг белого цвета, а ипаток – желтого. В 1988–1990 гг. на острове Талан ипаток было примерно в 1.5 раза меньше, чем сейчас, а конюг – в 3–4 раза больше (Андреев и др., 2010). Т.е. отношение численности ипатки к численности большой конюги увеличилось примерно в 5 раз, поэтому желтый помет стал встречаться примерно в 5 раз чаще. Получилось так, что в течение довольно короткого промежутка времени изменились некоторые цветовые предпочтения больших конюг. Причиной этого, видимо, являлось изменение фона колонии: желтый цвет стал более распространенным. Большие конюги в прошлом на искусственные объекты желтого цвета реагировали слабо, а в настоящее время реагируют гораздо сильнее.
Однако оранжевый цвет задал загадку. У конюг ярко-оранжевый клюв, поэтому, казалось бы, реакция на этот цвет должна быть выраженной. Тем не менее в июне, когда клювы больших конюг оранжевые, реакция на предметы этого цвета была слабой. С начала июля у больших конюг начинается линька рамфотеки, и оранжевые щитки с клюва начинают опадать. Почему-то именно в это время реакция больших конюг на оранжевые предметы резко усиливалась. Опыты 1989 г. проводились после 1 июля (Зубакин, Зубакина, 1990), чем и объясняется, на наш взгляд, повышенная реакция на оранжевый в тех опытах. Объяснить подобное изменение реакции на оранжевый цвет по ходу сезона пока не удалось.
Наименее предпочитаемым оказался красный цвет. У самок, однако, реакция на него была выражена достоверно сильнее, чем у самцов (таблица). Это, возможно, объясняется тем, что красный цвет ближе к оранжевому, чем к зеленому или синему, а на оранжевый у самок реакция тоже была сильнее, чем у самцов, причем после 1 июля эта разница становилась достоверной (таблица).
“Сложные” предметы предпочитались “простым”. Такая дифференцированность и такие предпочтения говорят о неутилитарности реакции выбора предметов большими конюгами. До сих пор считалось, что: “у животных с внесезонным размножением есть “избыточная” энергия, которая может тратиться на исследование обстановки и ее деталей, на манипулирование, на игровую деятельность и … другие формы активности” (Алексеева, 1977). Для животных с сезонным размножением считалось, что вся энергия уходит на размножение.
Во многих случаях это действительно так. Однако результаты данного исследования показали, что некоторые виды с сезонным размножением, но живущие плотными сообществами, проявляют исследовательскую активность даже в сезон размножения. Некоторые такие виды проявляют ярко выраженное любопытство и готовы отвлекаться от забот о размножении и социальной активности ради пусть и примитивного, но все же познания окружающего мира (Харитонов, 2006).
Примечательно, что такое поведение люриков и больших конюг напоминает манипуляционно-исследовательское поведение врановых птиц (Зорина и др., 1986), умственные возможности которых выше, чем у большинства других птиц. Однако по уровню развития мозга (отношение веса полушарий головного мозга к весу его стволовой части – индекс Портмана (Portman, 1947) чистиковые птицы (правда, среди чистиковых в той публикации рассмотрен только тупик) оказались примерно в 2 раза ниже, чем врановые.
Действительно, манипуляционно-исследовательское поведение отмечено у птиц с разным уровнем развития мозга (Дьюсбери, 1981), однако в разной степени. Большинство видов морских птиц реагировали на искусственные предметы, но только после того, как экспозиция этих предметов была гораздо продолжительнее, чем в опытах с большими конюгами и люриками. Ни у какого другого вида, кроме как у больших конюг, не отмечена реакция на цветы или на кольца на ногах своих соседей по колонии. Не исключено, что по степени направленности своего поведения на исследование предметов и времени манипулирования, большие конюги и люрики сопоставимы с врановыми птицами, хотя относительный размер мозга у чистиковых меньше.
При этом у обоих упомянутых видов чистиковых обращают на себя внимание высокая социальная активность, продолжающаяся весь гнездовой сезон, и сложная социальная структура (Харитонов, 2011). У других видов птиц и структура колонии, и интерес к окружающему миру, выходящие за рамки потребностей размножения, значительно меньше. Причина высокой и социальной, и исследовательской активности люриков и больших конюг не ясна, однако, не исключено, что корень этой активности один и тот же, например, соответствующий гормональный уровень, который пока совершенно не исследован.
Интерес к предметам стимулирует у морских птиц исследовательскую активность вообще. Видимо, люрики, конюги и ипатки, которые обратили внимание на необычные предметы, тут же демонстрировали всплеск общей исследовательской активности. Они начинали гораздо пристальнее, чем обычно, осматривать участки поверхности камней, цвет которых отличался от цвета фона, заглядывали за камни, трогали пятнышки на камнях, которые до этого никогда не трогали, теребили травинки и прочие мелкие естественные объекты.
Искусственные предметы и социальные отношения.
Люрики выстилают дно своей гнездовой камеры мелкими плоскими камешками. Мы неоднократно наблюдали, как птица передавала принесенный камешек партнеру другого пола и демонстрировала их другим люрикам. По нашим наблюдениям, точно так же люрики могли демонстрировать другим особям и предметы из опытов (рис. 4).

Рис. 4. Люрик предъявляет игрушечный подсвечник другому люрику.
Большие конюги и в меньшей степени люрики иногда использовали предметы как еще один инструмент выстраивания социальных отношений. У конюг это выражалось в том, что они начинают охранять эти предметы от других особей, хотя не всегда успешно. Например, если кто-то из группы конюг тронул клювом один из предметов, то другая особь из группы могла отогнать тронувшую не только от этого, но и от всех предметов. Люрики, если и охраняли предметы от других особей, то этого им было явно мало. Надо было еще и подтвердить каждый раз свой статус: если кто-то тронул какой-то предмет, например зеленый, то его могли не просто отогнать от этого предмета, но прогнавшая особь обязательно сама затем трогала этот зеленый предмет и далее охраняла его от возможности прикоснуться к нему других птиц. Если же другая птица трогала теперь белый предмет, то отгоняли именно от белого, и отгоняющая птица начинала трогать белый.
“Охрана” искусственных предметов у птиц, видимо, подобна охране тех естественных предметов, что птицы приобрели. В 1988 г. на о-ве Талан мы наблюдали, как один из топориков прилетел с пучком травы в клюве и долго стоял на одном месте. Потом бросил этот пучок на камень и примерно 5 мин охранял, демонстрируя угрожающие позы близко проходящим топорикам. Потом вдруг отошел от этого пучка травы, потеряв интерес к нему.
О том, что через отношение к посторонним предметам животные могут проявить и даже поменять свой социальный статус, свидетельствуют наблюдения за шимпанзе в природе. Пример такого влияния постороннего предмета на статус есть то самое наблюдение Джейн Гудолл, где молодой самец стучал по пустой канистре, чем вводил в ступор вожака (Гудолл, 1992). Демонстрация люриками предметов другим особям, возможно, повышает статус демонстрирующего.
Сходство манипуляционного поведения и игры.
У больших конюг и люриков хорошо видно наличие избытка энергии, которую, видимо, требуется куда-то использовать. В значительной мере энергия тратится на социальную активность. Однако, как оказалось, часть энергии может уходить и на манипуляционно-исследовательскую активность, которая во многом напоминает игру. Если предметы (“игрушки”) не очень яркие и небольшие, то люриков и больших конюг прежде всего привлекают знакомые цвета. Если же предмет большой и яркий, то более привлекательным становится он, даже если его цвет сильно отличается от привычного.
По нашему мнению, поведение конюг и люриков с предметами сильно напоминает поведение детей, переключившихся на новую игрушку или игру. Если птица (люрики, конюги, ипатки, топорики) трогает предмет, то тут же начинает трогать клювом пятна на камне (до этого внимания на них не обращала) и траву. Хватание птицами предметов, а также ощупывание пятен помета на камнях и травинок – это проявление явного любопытства с элементами игры. В этом плане максимальную в наших опытах игровую активность демонстрировала одна из ипаток, которая примерно 10 мин играла с двумя красными дисками. Она их хватала клювом, приподнимала, перемещала, откидывала. Один из дисков вскоре уронила с камня, затем долго смотрела, куда он упал, и продолжила манипуляции уже с одним оставшимся диском. Следует отметить, что у больших конюг элементы игры проявляются и в повседневном поведении, например молодые конюги могут устраивать турниры, затеяв “борьбу” шеями. Поэтому игра с посторонними предметами не выглядит совсем уж необычным явлением.
Поведение, свидетельствующее о возможном анализе птицами свойств предметов.
Основная часть наших данных и последующий анализ регистрирует и сравнивает только факт манипулирования необычными для птиц искусственными объектами. Однако некоторые наши наблюдения позволяют несколько глубже оценить структуру их исследовательского поведения. В ряде опытов мы наблюдали попытки птиц манипулировать естественными предметами непосредственно после касания предметов искусственных или манипуляции ими (хотя до манипулирования искусственными предметами на естественные объекты птицы внимания не обращали). По нашему мнению, таким способом происходило более глубокое исследование предметов. Это можно считать попытками птиц анализировать свойства искусственных предметов, сравнив их с естественными. Во время опытов ипатки иногда делали странные движения, которые не наблюдались у птиц, не интересовавшихся предметами. Ипатка в опыте 15.07.2008, стоя возле большого диска, несколько раз трогала камень, который находился на расстоянии примерно 20 см от этого диска. Она, возможно, “анализировала” и “сравнивала” камень и предмет.
Возможно, что ипатка может исследовать предмет, сопоставляя его со своим телом: она хватает не только черную точку на камне – но и, потрогав диск, несколько раз хватает себя за цевку. Вне этих опытов хватания клювом своей цевки (а не другой особи, как это отмечалось в брачном поведении (Харитонов, 1980)) у ипатки ни разу не наблюдалось. Интерпретировать такое поведение можно так: цвет ноги похож на цвет предмета, и если ощупали клювом предмет, надо ощупать и свою цевку. Птица, возможно, пытается “понять”, что есть предмет и таким образом его исследует. Не исключено, что птица сравнивает цвет ноги с цветом диска, которые действительно очень похожи.
Микро-локальные особенности поведения.
По результатам опытов удалось заметить еще одно поразительное явление, которое можно назвать микро-локальными особенностями исследовательского поведения у больших конюг. Известно (хотя и слабо изучено), что поведение особей одного вида в разных географических районах может различаться радикально. Оказалось, что радикальные различия могут быть у локальных группировок животных, даже – у микро-локальных. На нашей площадке был камень, который “портил” результаты.
Именно на этом камне результаты опытов на птицах, которые там находились, оказались противоположными таковым на всех остальных окрестных камнях. Если из наборов скрепок (5 розовых +5 омедненных +5 неомедненных) на других камнях первыми выбирали розовые скрепки, то на “неправильном камне” – омедненные. Второй раз сильное отличие было в опытах с черной пластиковой полоской. Везде первая реакция была не на черную полоску, а на другие предметы, а на “неправильном камне” – на черную. Третий случай – столь же примечательный и сильно “испортивший” общий результат (из-за чего камень и стал называться “неправильным”) – выбор между оранжевыми заколками и малым диском. Самки сначала брали заколки, а первая и последующие реакции самцов – брать не заколки, как на других камнях, а малый красный диск.
Следует отметить, что подобные микро-локальные особенности поведения отмечаются нами у птиц не первый раз. При наблюдении за поведением выводков тихоокеанских черных казарок (Brants bernicla nigricans) в Дельте Юкона в 1993 г. у двух выводков черных казарок нами отмечены резкие отличия от поведения всех остальных выводков именно в одном микро-локальном районе (Kharitonov, 1997). Причину подобных отклонений мы ни тогда, ни сейчас понять не могли. В случае с черными казарками мы тогда посчитали, что влияют некие “особые факторы”, еще не выявленные нами. Видимо, действием неких особых факторов можно предварительно обозначить и причину микро-локальных поведенческих различий у больших конюг.
Другие виды морских птиц.
На выложенные на камни предметы другие виды реагировали значительно слабее больших конюг. Ипатки иногда брали предметы, но знакомых цветов – бордового, желтого и зеленого. Бордовый и желтый цвета присутствуют в окраске клюва ипаток, зеленый – похож на цвет травы. Ипатки трогали также красные диски, причем в одном случае ипатка увлеченно играла с двумя пластиковыми дисками.
У малых конюг и конюг-крошек реакция на предметы была сходна с таковой у ипаток. Эти два вида птиц не реагировали на мелкие цветные предметы, но активно и подолгу ощупывали клювом яркий красный диск, по размеру превосходящий мелкие предметы. У малых конюг при этом наблюдалась “наведенная” реакция, подобная той, что наблюдалась и у больших конюг: потрогав клювом яркий красный диск, некоторые малые конюги трогали клювом мелкий цветной предмет, на который в отсутствие более сильного раздражителя – красного диска – реакции не было.
Интерес к необычным предметам обнаружен также у глупышей (рис. 5). Эти птицы, даже занятые нуждами размножения, трогали и ощупывали клювом предъявленные им предметы. Однако, в отличие от чистиковых птиц, они довольно быстро теряли к таким предметам интерес.

Рис. 5. Глупыш ощупывает клювом большой красный диск.
Опыты на о-ве Ионы были нацелены, прежде всего, на малых конюг. При этом большие конюги стали трогать предметы клювом сразу вскоре после их закрепления на камне, хотя трогать их приходилось из очень неудобного для больших конюг положения тела, поскольку предметы были приклеены к поверхности наклоненного под большим углом камня. Для малых конюг (для этого вида такие наклонные камни являются основным биотопом) и конюг-крошек потребовалось, видимо, накопительное действие раздражителей. Приклеенные к камню скрепки и заколки эти птицы первые два дня не трогали. Один день не трогали они и помещенный на камень небольшой пластиковый красный диск диаметром 5 см. Только спустя сутки малые конюги и конюги-крошки стали трогать и теребить клювом более яркий предмет – красный диск; и только после него обратили внимание и стали трогать камень и цветную скрепку. Можно сказать, что малая конюга и конюга-крошка проявляют интерес к инородным предметам примерно на том же уровне, как и ипатки.
Восприятие запахов и вкуса.
В наших опытах у больших конюг не отмечено никакой реакции на запах ментола, хотя с точки зрения наблюдателя запах был чрезвычайно сильный и едва переносимый. Не отмечено нами также реакции на запах лимона и на запах (но не на вкус) чеснока. Проведенные нами опыты показали, что некоторые запахи, в частности, запах 10% раствора аммиака, судя по поведению, птицы воспринимают. Этот запах явно создавал конюгам некоторые неприятные ощущения: они задерживали клюв на пути к предметам и отдергивали его, прежде чем сделать повторные попытки взять предмет. Явно неприятные ощущения не являлись препятствием для схватывания предмета клювом, хотя предъявлявшиеся предметы с чисто утилитарной точки для конюг были бесполезны, неприятные ощущения от запаха аммиака их не останавливали. То же самое и с чесноком. Возможно, вкусовая и запаховая чувствительность у самцов в наших опытах была несколько выше, чем у самок. Однако, учитывая небольшую величину выборок, эти различия проявились лишь как тенденция.
После первых публикаций о том, что большие конюги в состоянии ощущать и различать запахи, многие исследователи стали преувеличивать способности птиц в использовании этой чувствительности. Появились даже воззрения, что конюги могут использовать запахи для социальных отношений и даже переносить запах друг на друга (Зубакин и др., 2010). Дальше гипотез здесь дело не пошло, подтверждающих сведений на эту тему не получено, однако нами получены сведения совсем другого рода.
Что касается запаховой сигнализации больших конюг, то представление о значительной поведенческой роли этого явления у нас давно вызывало сильный скепсис. Так, в 1988 г. на о-ве Талан мы проводили следующие эксперименты: из погибших птиц изготовляли макеты и выставляли их на снег для наблюдения за реакцией конспецификов.
Макеты изготовляли следующим образом: в погибшую птицу вводили две гибкие металлические проволоки, при помощи которых телу придавали ту или иную позу для имитации живой птицы. Чтобы предотвратить быструю порчу макета, в него при помощи шприца накачивали 4% раствор формальдегида, точно таким же образом мы ранее изготовляли макеты озерных чаек для проведения экспериментов по изучению привлекательности групп макетов, имитирующих живых птиц (Харитонов, 1984). Несмотря на то, что выставляемые на поверхность колонии макеты конюг сильно пахли формалином, а от некоторых уже шел заметный запах разложения, живые птицы положительно реагировали на макеты, приближались к ним, прижимались к ним шея к шее и погружали свой клюв в оперение макетов, как в живых птиц. Конюги “токовали” с макетами, даже когда они сильно испортились, т.е. неприятный запах им явно не мешал. Очевидно, что чужеродные запахи нисколько не смущали живых птиц, они реагировали на макеты как на чисто визуальные стимулы.
Подобное показали и эксперименты тех же американских ученых, которые открыли ольфакторную чувствительность у конюг. В T-образном лабиринте большие конюги достоверно чаще по сравнению с контролем выбирали: 1) перья больших конюг, 2) выделенное из перьев то самое пахнущее цитрусом вещество; 3) не демонстрировали предпочтения амил-ацетата (банановая эссенция); 4) достоверно негативно, по сравнению с контролем, реагировали на мускус млекопитающих (Hagelin, 2007; Hagelin et al., 2003).
Дальше теми же учеными были поставлены эксперименты для выяснения роли запахов в общении конюг. Было взято 12 макетов больших конюг: 6 самцов и 6 самок. По реакции приближения к макетам на колонии 555 самцов и 280 самок не найдено свидетельств сексуального предпочтения моделей с запахом по сравнению с моделями без запаха. Самки, которые подходили к моделям самцов с искусственно добавленным запахом, не с большей вероятностью исполняли половые демонстрации по сравнению с самками, которые приближались к контрольным моделям с более слабым запахом (Jones et al., 2004). Меньше самцов приближались к моделям самок без запаха, но эффект был тот же – сексуальное поведение одинаково по отношению к птицам с запахом и без запаха. Однако самцы приближались к моделям самцов с запахом ближе и на более длительное время, чем к контрольным моделям самцов. Самки также приближались ближе к пахнущим моделям самцов, при этом разницы в числе сексуальных демонстраций не было.
Совсем другое происходило, когда у конюг обрезали хохол. Число сексуальных демонстраций, направленных на самок с хохлом, было в два раза больше, чем на самок без хохла. На самцов с хохлом – в 4 раза больше, чем на самцов без хохла. Таким образом, в сексуальном поведении большие конюги больше полагались на зрение, чем запах (Jones et al., 2004).
Для люриков не известно наличия секреции каких-либо специфических запаховых веществ. Тем не менее, люрики тоже показали, что они в состоянии ощущать, по крайней мере, некоторые запахи. Люриков, которые имели намерение взять предметы, запах ментола от этих предметов останавливал; птицы в результате прекращали попытки дотронуться до таких предметов. Свидетельств того, что эти птицы используют запах в социальных отношениях, на настоящий момент нет. Птицы запаховой чувствительностью обладают, но, в отличие от зрения, которое используется всегда, могут воспользоваться обонянием, а могут – и нет (Э.П. Зинкевич, устное сообщение).
Показано, что птицы могут воспринимать и различать запахи, однако еще не доказана их способность различать по запаху отдельных особей, т. е. способность чувствовать индивидуальные запахи. Передача запаха от особи к особи (Зубакин и др., 2010) имеет смысл и имеет место у тех животных, которые способны различать запахи индивидов. У птиц данная способность не обнаружена, поэтому, на наш взгляд, нет оснований предполагать наличие передачи запаха от особи к особи у больших конюг. Кроме того, у больших конюг территориальная, а не семейно-пространственная структура поселений, и передача запаха по условиям их жизни, по нашему мнению, вовсе не требуется. Поэтому у ряда исследователей наступило некоторое разочарование относительно социальной роли запаховой сигнализации у конюг. Существуют и другие гипотезы о причинах выделения запаховых агентов у больших конюг. Например, защита от паразитов (Douglas et al., 2004), правда, иной раз, эти гипотезы причудливо переплетаются (Douglas, 2008).
Заключение
У морских птиц отмечен интерес к предметам окружающего мира, не имеющим отношения к поддержанию жизни и размножению. Птицы обращают повышенное внимание на выделяющиеся из фона предметы, пытаются их исследовать визуальным и тактильным способами. В наибольшей мере этот интерес отмечен у высокосоциальных морских птиц: люриков и больших конюг. Их манипуляционно-исследовательская активность выражена значительно сильнее, чем у других видов морских птиц. Социальная активность люриков и больших конюг тоже значительно выше, чем у других видов чистиковых птиц (Харитонов, 2011).
Возможно, что эти два вида активности происходят из некоего общего физиологического и поведенческого корня. В отдельных опытах удалось отметить использование птицами ольфакторных и вкусовых рецепторов.
В наших опытах имеются два необъяснимых результата:
1) усиление реакции больших конюг на оранжевый цвет в конце сезона размножения; это чаще наблюдается у самок, чем у самцов;
2) наличие буквально микроскопических по размеру мест, где поведение больших конюг резко отличалось от поведения птиц этого вида в других микро-местах.
Необходимо подчеркнуть, что по данной тематике получены только первые сведения. Однако они уже показали, насколько поведение морских птиц сложнее нашего представления о нем. Манипуляционно-исследовательская активность морских птиц, несомненно, требует дальнейшего более глубокого изучения.
Благодарности
Автор выражает искреннюю благодарность сотрудникам Института биологических проблем севера ДВО РАН (г. Магадан) А.Я. Кондратьеву, А.С. Китайскому, А.В. Андрееву, Е.В. Голубовой, а также сотруднику ИПЭЭ РАН В.А. Зубакину, сотруднику Полярного Института, г. Тромсё, Норвегия Хальвару Стрёму (Hallvard Strom), участнику экспедиций на о-в Медвежий, Шпицберген, С.А. Чельцову. Без их помощи и поддержки данное исследование едва ли было возможно. Огромную благодарность автор выражает З.А. Зориной, докт. биол. наук, профессору, зав. лабораторией физиологией и генетики поведения биологического факультета МГУ им. М.В. Ломоносова за глубокое обсуждение материала и весьма ценные замечания, радикально улучшившие статью.
Список литературы
Алексеева Л.В., 1977. Полицикличность размножения приматов и антропогенез. М.: Московское общество испытателей природы. 105 c.
Андреев А.В., Голубова Е.Ю., Зубакин В.А., Харитонов С.П., 2010. Численность морских птиц на колониях о. Талан: двадцатилетний тренд // Вестник СВНЦ ДВО РАН. No 2. С. 30–42.
Вааль, Франс де, 2017. Достаточно ли мы умны, чтобы судить об уме животных? М. “АНФ”. 404 с.
Виноградов А.А., 2015. Опыт определения пола люриков (Alle alle Link) по очертаниям клюва // Самарский научный вестник. No 2 (11). С. 47–51.
Гудолл Дж., 1992. Шимпанзе в природе: поведение. М.: Мир. 670 с.
Дерягина М.А., 1986. Манипуляционная активность приматов. М.: Наука. 110 с.
Дьюсбери Д.М., 1981. Поведение животных. Сравнительные аспекты. М.: Мир. 480 с.
Зорина З.А., Маркина Н.В., Дерягина М.А., 1986. Структура и возрастные особенности манипуляционной активности серой вороны (Corvus cornix) // Зоологический журнал. Т. 65. No 10. С. 1552–1558.
Зубакин В.А., Володин И.А., Клёнова А.В., Зубакина Е.В., Володина Е.В. и др., 2010. Поведение большой конюги (Aethia cristatella) в период размножения: двигательные и акустические демонстрации // Зоологический журнал. Т. 89. No 3. С. 331–345.
Зубакин В.А., Зубакина Е.В., 1990. Об “эстетических вкусах” больших конюг // Изучение морских колониальных птиц в СССР. Информационные материалы. Магадан: ИБПС. С. 39–42.
Конюхов Н.Б., 1996. Морфо-экологические и этологические различия в семействе чистиковых (Aves: Alcidae) // Известия РАН. Серия биологическая No 4. С. 446–452.
Лакин Г.Ф., 1990. Биометрия. М.: Высшая школа. 343 с.
Обозова Т.А., Багоцкая М.С., Смирнова А.А., Зорина З.А., 2013. Сравнительная оценка способности разных видов птиц к решению протоорудийных задач // Зоологический журнал. Т. 92. No 6. С. 1–11.
Обозова Т.А., Смирнова А.А., Артюхин Ю.Б., 2010. Экспериментальный подход к изучению когнитивных способностей серокрылых чаек (Larus glaucescens) в их естественной среде обитания: относительные оценки размера предметов, обучение путем наблюдения за конспецификами // Орнитология в Северной Евразии. Материалы XIII Международной орнитологической конференции Северной Евразии. (Оренбург, 30 апреля – 6 мая 2010). Оренбург: Издательство Оренбургского государственного педагогического университета. С. 237–238.
Плохинский Н.А., 1978. Математические методы в биологии. М.: МГУ. 266 с.
Поповкина А.Б., Барсова Л.И., Анисимов В.Д., 2010. Изучение обоняния птиц в России и за рубежом: результаты, проблемы и перспективы // Орнитология в Северной Евразии. Материалы XIII Международной орнитологической конференции Северной Евразии.
(Оренбург, 30 апреля–6 мая 2010). Оренбург: Издательство Оренбургского государственного педагогического университета. С. 1254–1255.
Резникова Ж.И., 2005. Интеллект и язык животных и человека. Основы когнитивной этологии. М.: Академкнига. 520 с.
Харитонов С.П., 1980. Материалы по птицам острова Ионы // Орнитология. Вып. 15. М.: МГУ. С. 10–15.
Харитонов С.П., 1984. Изучение сигнального значения макетов в ориентации озерных чаек // Сигнализация и экология млекопитающих и птиц. М.: Наука. С. 235–246.
Харитонов С.П., 2006. Пространственно-этологическая структура колоний околоводных птиц. Автореф. дис. … докт. биол. наук. М.: ИПЭЭ РАН. 48 с.
Харитонов С.П., 2011. Пространственно-этологическая структура колоний околоводных птиц // Зоологический журнал. Т. 90. No 7. C. 846–860.
Cramp S., 1985. The birds of the Western Palearctic. Oxford: Oxford University Press. V. 4. 960 p.
Douglas H.D., 2008. Prenuptial perfume: all anointing in the social rituals of crested auklet (Aethia cristatella) and the transfer of arthropod deterrents // Naturwissenschaften. V. 95. P. 45–53.
Douglas H.D., Co J.E., Jones T.H., Conner W.E., 2004. Interspecific differences in Aethya spp. auklet odorants and evidence for chemical defense against ectoparasites // Journal of Chemical Ecology. V. 30. No 10. P. 1921–1935.
Hagelin J.C., 2007. The citrus-like scent of crested auklets: reviewing the evidence for an avian olfactory ornament // Journal of Ornithology. V. 148 (Suppl. 2). P. S195–S201.
Hagelin J.C., Jones I.L., Rasmussen L.E.L., 2003. A tangerine-scented social odour in a monogamous seabird // Proceedings of the Royal Society of London. Series B. 270. P. 1323–1329.
Jones I.L., Hagelin J.C., Major H.L., Rasmussen L.E.L., 2004. An experimental field study of the function of crested auklet feather odor // Condor. V. 106. P. 71–78.
Kharitonov S.P., 1997. Brood interactions in the Pacific Black Brant (Branta bernicla nigricans). Casarca. Bulletin of Goose and Swan Study Group of Eastern Europe and North Asia. Publ. М.: Издательство Россельхозакадемии. No 3. С. 138–155.
Portmann A., 1947. Etudes sur la cerebralisation chez les oiseaux. II. Les indeces intra-cerebraux // Alauda, V. 15. No 1. P. 1–15.
Stempniewicz L., 1986. Factors causing changes in the rhythm of attendance in the little auks, Plautus alle (L.) at a colony during the breeding season in Svalbard // Ecology Polska. V. 34. No 2. P. 247–263.
Зоологический журнал. Т.97. №4.
Свежие комментарии