 Print
Print  PDF
PDF
Рис. 1. «Хотелось лечь, прикрыть бы телом…», — пел Леонид Утесов. На картинке видно, как божья коровка прикрывает телом вышедшую из нее личинку наездника. Яркая окраска отпугивает хищников, а для большего устрашения насекомое периодически подрагивает всем телом. Так продолжается примерно неделю, после чего наездник выходит из яйца и улетает, а божья коровка… Божья коровка даже иногда выживает, но это неважно, неважно. Источник изображения – Maure et al., 2011
Резюме. Публикуется обзор М.В.Гопко и В.Н.Михеева, посвящённый паразитическим манипуляциям — направленным изменениям паразитом морфологии и поведения хозяина, повышающим приспособленность первого, с интересными и новыми данными о паразитах как «клее» биотических отношений в экосистемах, с критическими замечаниями публикатора.
От публикатора: что не так с «парадигмой Панглосса» применительно к паразитическим манипуляциям?
Содержание
Последние вызваны некритическим (сорри за тавтологию) следованиям идеям Р.Докинза и, шире, социобиологии, рассматривающей тела индивидов, реализующие «должное» поведение в «должных» моментах взаимодействия, как управляемые изнутри комбинациями «эгоистичных генов» (Alexander, 1974, 1975). Последние неизбежно додумываются, выступая в лучшем случае полезными фикциями; современные данные об организации генных сетей, их участия в «эволюции в пространстве возможностей» показывают что
а) гены — никак не «эгоистичные репликаторы» Р.Докинза, они иерархически организованы и управляемы «сверху», со стороны организма, как любые другие морфоструктуры;
б) именно организмы, не отдельные гены, оказываются единицей отбора на уровне популяции. Последний же совершенствует не индивидов с их качествами, выживанием или размножением (все эти параметры могут и ухудшаться, как падает размножаемость по мере прогресса социальности у грызунов), но разного рода приспособления вида, от морфологии до жизненных стратегий, в т.ч. описанные в статье паразитические манипуляции. Сюда же относятся регуляторы, выступающие «передаточным механизмом» в генных сетях, от сигналов об изменении внешней/внутренней среды, до компенсирующих ответов в виде изменений экспрессии генов, что в эволюционных масштабах времени меняет темп эволюции как самих генетических «текстов», так и «конструкции» генных сетей, «читающих» их для оптимального реагирования на изменения среды, как фабрики отвечают на изменения рыночной конъюнктуры. Сами индивиды, их перемещения и взаимодействия друг с другом ли, с биоценотическими ли партнёрами, оказываются лишь «статистическими испытаниями» и «расходным материалом» для вышеописанного (это т.н. социоцентрический — или популяционноцентрический взгляд на поведение);
в) вопреки всем заявлениям социобиологов, «эгоистичные репликаторы», «гены альтруизма/эгоизма», вообще «гены поведения» и т.д. мысленные конструкты до сих пор не связали с конкретными генными вариантами, и не особо пытаются. Поскольку картина реальной эволюции соотносится с социобиологическим описанием происходящего как грани куба Неккера (используя замечательную метафору из «Эгоистичного гена»). Если преобразования тел организмов, в т.ч. видоспецифических форм поведения, обслуживающих меняющиеся отношения особей в популяции, есть главное в эволюции, то изменения на уровне генов и/или физиологии, связанных с первыми и влияющих на них – их «тень» (эпифеномен) или, точнее «отдача» происходящей эволюции. Организация меняется в ответ на эти последние так, чтобы обеспечить устойчивость новшеств, поддерживаемых и усиливаемых отбором, и при этом не вымереть (поскольку и организм, и его генетические системы целостны).
И действительно, в настоящей статье много и интересно рассказывается о поведенческих и/или физиологических феноменов, связанных с манипуляцией хозяином, и реакцией последнего на эти воздействия и совсем ничего — о генах того и другого, «эгоистически» увеличивших своё реплицирование благодаря подобным приёмам. А ведь согласно идеологии автора, с этого надо б начать!
На деле сколько-нибудь подробный анализ показывает, что «гены», «эгоистичность» и «отбор на уровне генов» здесь — лишь риторические приёмы, дающие видимость объяснения. Неслучайно среди исследований паразитических манипуляций, фигурирующих в настоящем обзоре, во многих реконструированы проксимальные механизмы, обеспечивающие их эффективность и биоценотическую роль благодаря «ближайшим эффектам» поведенческих воздействий паразита и физиологических реакций на них хозяина, но нет ни одной реконструкции ультимаативных механизмов, выводящих наблюдаемые в природе манипуляции из вышеназванной «докинзовской триады».
«Эгоистичность» понимается как управление геном «подвластных» ему аспектов поведения организма, но не только его – также и биоценотических партнёров (Hamilton, 1963, 1964, 1971; Trivers, 1971, 1972, 1974). Это т.н. «длинная рука гена», одно из проявлений которой — обсуждаемые паразитические манипуляции. Последние, как и «управление» телами самих организмов, по мысли социобиологов, производится генами так, чтобы максимизировать их репликацию и передачу в нисходящем колене, даже когда это вредит выживанию (затрудняет существование) тел, самого ли животного, или его биоценотических партнёров.
Эволюция выступает здесь как «слепой часовщик». Она подбирает нужные организму паттерны среды и параметры стимуляции а) «кусок за куском», как бы собирая паззл, б) «вслепую», через попытки управлять ими, реализуя определённое поведение организмов, и отбор успешных попыток) так, чтобы «на финише» жизненного цикла особей, после реализации всего поведения, во всех видотипичных жизненных обстоятельствах, обеспечивать максимально расширенное воспроизводство копий данного репликатора (гена или генного варианта) в сравнении с конкурирующими вариантами (Wilson, 1975; Докинз, 1993, 2006).
По мысли социобиологов, «эгоизм» состоит в усилении и качественном «оттачивании» этого управления отбором на уровне генов (а не фенотипа), естественно, если «выигрыш» от манипуляции превзойдёт «плату». Это крупнейшее методическое достижение социобиологии — мысль, что эволюционное новшество в морфологии ли, в поведении ли, помимо выигрыша (benefit), следующего из адаптивности, всегда предполагает «плату» (cost) в виде 1) затрат вещества, времени и энергии на образование соответствующих форм и структур, 2) специфических рисков при «должном» предъявлении данных органов, например, брачных украшений и ритуализированных демонстраций, в «должной» ситуации взаимодействия, почему 3) Отбор формирует соответствующую манипуляцию — что коммуникативную, посредством эффектов видовых демонстраций (Dawkins, Krebs, 1978), что паразитические, анализируемые в данной статье, лишь в том случае, когда «выигрыш» от неё существенно превосходит «плату» (измеряемую в конечном счёте не в затратах энергии или пластических веществ, а в последствиях их совершения для итоговой приспособленности). В ситуации «за морем телушка полушка, да рупь перевоз» манипуляции не формируются, какие б не следовали выгоды от появления «у гена» такой «длинной руки».
Здесь проявляется ещё одно противоречие социобиологии — между методическими достижениями и дефектной теорией, толкающей к т.н. эффекту Панглосса, когда каждая хорошо выраженная особенность биологии вида представляется адаптацией, «для чего-то полезной» и «чем-то выгодной». Внутренние противоречия этого взгляда и следующие из него ошибки разобраны в знаменитой статье Ст.Дж.Гулда и Р.С.Левонтина про «Пазухи свода собора святого Марка и парадигму Панглосса» («Spandrels…, 1979), и я лишь добавлю ещё одно.
Он превращает науку в область счастливых догадок (как высказался А.Куприянов о «популяризации» этологии в известных книгах В.Р.Дольника, однако, увы, это в полной мере присуще и «настоящей науке»). Какой бы паттерн поведения ни остановил на себе взгляд исследователя, всё оказывается адаптивным, надо лишь догадаться почему (чтоб потом подобрать данные «под гипотезу»), хотя есть более чем убедительные основания считать, что все без исключения первые впечатления такого рода будут ошибочными — даже о структуре поведения (членение на единицы, их форма и пр.) , а уж тем более об их функции, см. «Уязвимость «парадигмы Панглосса-1, -2, -3″.. К тому же догадки высказываются от имени «современной (читай — единственно верной) эволюционной теории», толкая её авторитетом соответствующие объяснения всячески подкреплять и «спасать», когда плохо соответствуют фактам, вместо необходимого для учёного трезвого скепсиса в первую очередь к следствиям собственной концепции, теоретическим или проверенным в опыте, ведь без этого её не усовершенствуешь, здесь попперовский фальсификационизм рулит.
***
М.В.Гопко, В.Н.Михеев
Паразиты воздействуют на своих хозяев самыми различными способами. В тех случаях, когда они меняют фенотип своих хозяев с целью увеличить собственную приспособленность, говорят о манипуляциях паразитов фенотипом хозяина. Вероятно, не все паразиты манипулируют поведением своих хозяев, но, несомненно, паразитические манипуляции весьма распространенный природный феномен. Паразит может менять фенотип хозяина, чтобы обеспечить себе дополнительную защиту, создать оптимальные условия для роста и развития и, разумеется, чтобы успешно решить задачу переноса от одного хозяина к другому. Стратегии манипулирования необычайно многообразны и тесно связаны с жизненным циклом паразита. В обзоре мы даем представление об основных типах манипулятивных стратегий и обсуждаем, как та или иная стратегия могла сформироваться в ходе эволюции, почему одни стратегии встречаются чаще, чем другие, и как меняется стратегия манипулирования по ходу роста и развития паразита. Только недавно стали осознавать, что паразитические манипуляции — фактор, влияющий как на взаимоотношения в инфрасообществе паразитов внутри хозяина, так и на всю экосистему в целом. В двух последних разделах обзора мы рассказываем о конфликтах и кооперации между паразитами-манипуляторами, а также о роли паразитических манипуляций в экосистемах. Завершается обзор мысленным путешествием в недалекое будущее, в ходе которого мы коротко рассказываем о том, какой может стать роль паразитов и их манипуляций в этом изменяющемся на глазах мире.
Вниманию читателей предлагается обзор, посвященный манипуляциям паразитов. Под паразитическими манипуляциями понимают изменения фенотипа хозяина, вызванные паразитом и повышающие приспособленность последнего (Dawkins, 1999; Poulin, 2010). Мы старались охватить тему максимально широко, чтобы показать, как паразитические манипуляции могут быть связаны с другими актуальными темами современной биологии.
Нам кажется, что это отражает современные тенденции в этой отрасли знаний, поскольку стало очевидно, что паразитические манипуляции это не забавный эволюционный курьез, а широко распространенный феномен, имеющий большое значение в живой природе (Dawkins, 2004, Poulin, 2010; Moore, 2013; Hughes, 2014; Heil, 2016). Тем не менее, многие важные вопросы, например манипуляции паразитов, переносящихся половым путем (Heil, 2016), роль манипуляций в экосистемах и пр. до сих пор изучены очень мало (Poulin, 1999; Lefevre et al., 2009; Weinersmith, Faulkes, 2014).
Ситуация осложняется еще и тем, что большинство паразитов одновременно существуют в двух экосистемах: внутренней — в организме хозяина, где им приходится взаимодействовать с другими паразитами, и внешней, где обитает хозяин и свободноживущие стадии паразита (Poulin, 1999). Кроме того, стратегии манипулирования поведением хозяина, применяемые паразитами, необычайно многообразны, как и возможные взаимодействия между паразитами и экосистемами, в которых они обитают. В нашем обзоре мы пытались дать представление как о достаточно глубоко изученных темах, так и о самом переднем крае науки о паразитических манипуляциях. Новых интересных направлений в изучении манипуляций сейчас формируется великое множество, и мы не претендуем на то, что наш обзор окажется всеобъемлющим. Мы, скорее, пытались подробно описать направления, казавшиеся нам наиболее интересными и перспективными, обращая внимание в первую очередь на самые современные и цитируемые исследования. Хотя паразитические манипуляции представляют большой теоретический интерес для эволюционистов (теория даже немного опережает экспериментальные исследования (Poulin, Maure, 2015)), мы по мере возможности старались иллюстрировать каждое новое понятие или гипотезу примерами из экспериментальных исследований, не вдаваясь в излишние отвлеченности.
Обзор состоит из четырех частей. В первой мы обсуждаем некоторые терминологические тонкости, даем краткий очерк развития представлений о паразитических манипуляциях и подробно обсуждаем место манипуляций в современных представлениях об эволюции живых систем. Во второй части мы описываем основные типы манипуляций и делаем попытку их классифицировать. Эта классификация в достаточной степени условна, но мы надеемся, что она поможет читателю уложить в голове большое количество представленных нами фактических данных. В третьей части обзора мы рассказываем о манипуляциях неинвазионных паразитов и касаемся вопросов, связанных с обитанием паразита во внутренней экосистеме — организме хозяина: конфликтов и кооперации между паразитами, а также затрат на манипуляции. Наконец, в последнем разделе мы обсуждаем, какие воздействия может оказывать паразит на экосистему в целом.
I. Паразитические манипуляции: терминология, история, теоретические предпосылки и перспективы
Манипуляции паразитов фенотипом vs манипуляции поведением хозяина.
Одной из центральных задач паразитологии является изучение способов переноса паразитов или их потомства от одного хозяина к другому (Thomas et al., 2005). Среди большого количества возможных стратегий есть одна, которая вот уже на притяжении нескольких десятилетий привлекает интерес исследователей и вызывает оживленную дискуссию,— стратегия манипуляции паразита фенотипом своего хозяина. Традиционно принято говорить о манипуляциях паразита поведением хозяина, однако, строго говоря, манипуляции нередко затрагивают целый комплекс фенотипических черт хозяина как поведенческих, так и морфологических, и физиологических. Например, трематоды Leucochloridium spp. поселяются в глазных щупальцах улиток, окрашивая их в яркие цвета и заставляя пульсировать (Dawkins, 1999; Wesolowska, Wesolowski, 2013).

Янтарка обыкновенная, поражённая трематодой Leucochloridium paradoxum
Подобные изменения фенотипа улитки делают ее более заметной для следующего хозяина паразита — птицы. По всей видимости, птицы принимают ярко окрашенные щупальца улитки за гусениц или каких-то других личинок насекомых, склевывают их и в свою очередь заражаются трематодами, помогая паразиту завершить жизненный цикл (Dawkins, 1999: Пер. с англ. Докинз, 2010; Wesolowska, Wesolowski, 2013). Изменения во внешнем виде улитки сопровождаются изменениями в ее поведении: зараженные спороцистами Leucochloridium spp. улитки более активны, совершают длинные переходы, проводя много времени на открытых пространствах и поднимаются выше по стеблям растений по сравнению с незараженными улитками (Wesolowska, Wesolowski, 2013). Хотя в данном случае влияние паразита на своего хозяина не ограничивалось поведением, воздействие Leucochloridium spp. на фенотип улитки тоже принято называть паразитической манипуляцией (Wesolowska, Wesolowski, 2013).
То же относится, например, и к нематодам Myrmeconema neotropicum, паразитирующим в абдомене муравьев Cephalotes atratus (Yanoviak et al., 2008). Если незараженные муравьи полностью окрашены в черный цвет, то брюшко зараженных ярко-красное и приподнято вверх. В этом примере паразиты вновь сочетают манипуляцию поведением с манипуляцией внешним обликом хозяина. Вероятно, ярко окрашенное брюшко муравья привлекает птиц, принимающих его за ягоду, они съедают муравья и разносят яйца нематод со своими фекалиями (Yanoviak et al., 2008). Еще одним примером не поведенческих манипуляций паразита может служить феминизация самцов или индукция партеногенеза вольбахией (Volbachia spp.) (Werren et al., 2008) — бактерией, которая является облигатным паразитом яйцеклеток многих видов беспозвоночных (например, от 40 до 66% наземных членистоногих по современным оценкам заражены этим паразитом (Zug, Hammerstein, 2012).

Муравей Cephalotes atratus, заражённый нематодой Myrmeconema neotropicum, поднимает покрасневшее брюшко.
Развитие представлений о манипуляциях паразитов фенотипом хозяина.
Первые предположения о том, что паразиты способны менять фенотип своего хозяина с целью увеличить собственную приспособленность появились еще в середине XX столетия. Например, Доббен (Dobben, 1952) обратил внимание на тот факт, что рыбы, зараженные цестодой Ligula intestinalis, значительно чаще становятся жертвой рыбоядных птиц — окончательных хозяев этого паразита. Однако первыми, кто начал рассматривать изменения поведения хозяина с точки зрения адаптации паразита, были канадские исследователи Бетл и Холмс, выпустившие в середине 1970-х годов серию статей, посвященных этому вопросу (Bethel, Holmes, 1973, 1974, 1977). Они изучали поведение амфипод (Gammarus lacustris и Hyalella azteca), зараженных скребнями (несколько видов), окончательные хозяева которых — водоплавающие птицы. Незараженные рачки — фотофобы, при малейшей угрозе ныряющие на глубину. Однако амфиподы, зараженные цистакантами скребней, ведут себя иначе. Во-первых, в отличие от незараженных рачков, они фотофильны и стремятся к свету. Во-вторых, почувствовав опасность, они не ныряют а, напротив, скользят в верхних слоях воды и/или прикрепляются к мелкому мусору на поверхности водоема (Bethel, Holmes, 1973, 1974). Более того, поведение рачков менялось именно тогда, когда паразиты достигали инвазионной стадии, т.е. были готовы заразить следующего хозяина (Bethel, Holmes, 1974). Наконец, Бетлу и Холмсу удалось показать, что эти поведенческие изменения действительно увеличивают вероятность попадания амфипод в пищеварительный тракт водоплавающей птицы (Bethel, Holmes, 1977). Все эти данные позволили этим ученым высказать предположение, что паразит в их экспериментах целенаправленно менял поведение своего хозяина с целью увеличить собственную приспособленность. Подобный подход привлек внимание широкого круга исследователей-экспериментаторов, и к настоящему моменту число примеров паразитических манипуляций уже исчисляется сотнями (Poulin, 2010).
Опыты В. Бетла и Дж. Холмса могли бы остаться незамеченными, не совпади они по времени с глобальной сменой парадигмы в биологии. В 70-е годы XX века взгляд биологов на организм, как таковой, претерпел существенные изменения. Во-первых, биологи обратили внимание, что естественный отбор, на самом деле, действует не на организменном уровне, а на уровне генов (Dawkins, 1976: Пер. с англ. Докинз, 1993). Организм же представляет для генов всего лишь удобную среду обитания или, как метафорически выразился известный эволюционист Р. Докинз,— “машину выживания”. Во-вторых, на поведение животных (как, впрочем, и на их фенотип в целом) оказывают влияние самые различные гены, зачастую находящиеся в разных организмах (Докинз, 2010). Простейший пример, пение самца канарейки заставляет клетки гипофиза самки производить больше гонадотропинов. Другими словами, гены самца, ответственные за пение, “коварно вмешиваются” в работу гипофиза самки, чтобы подготовить ее к спариванию (Докинз, 2010, с. 117— 118). В-третьих, интересы генов нередко расходятся, и тогда фенотип особи является некоторым “компромиссом” между двумя крайностями (выгодой только для одной или только для другой группы генов)(Докинз, 2010, гл. 8,13). Эти соображения легли в основу концепции “расширенного фенотипа”, которую можно кратко сформулировать так:
“поведение животного стремится максимизировать выживание “генов этого поведения” независимо от того, находятся ли гены этого поведения в организме животного, данное поведение осуществляющего” (Докинз, 2010, с. 393).
[Здесь надо продолжить «…только в том случае, если выживание «эгоистичных генов» посредством данного способа (морфологии, биохимии, поведения — той же манипуляции) будет «прибыльней» выживания посредством всех прочих возможных способов, и в первую очередь способа «ничего не менять». Ведь кроме «выигрышей» (benefit), ожидаемых от приспособлений вроде манипуляции, их выработка отбором с т.з. социобиологов предполагает «плату», связанную, например, с дополнительной элиминацией особей, снижающих эффективную численность популяции, с затратами на перестройку физиологии-биохимии у «лучше манипулирующего» паразита и пр. (cost).
И именно с т.з. социобиологии «выигрыш» от выживания через данное поведенческое изменение должен превзойти «плату», чтобы «включить» вырабатывающий его отбор. Или, точней, отношение «выигрыша» к «плате» у данного поведенческого приспособления выше, чем у его возможных альтернатив, включая тот вариант поведения, что уже присутствует в популяции до начала отбора. Т.е. отбор «включается» лишь для выработки «самых прибыльных приспособлений», а не любых возможных. Это — крупнейшее методическое достижение социобиологии, позволившее сравнивать поведенческие альтернативы, жизненные стратегии, и т.д. «точки приложения» и продукты отбора в своего рода единой валюте «прибыльности в итоговой приспособленности» (inclusive fitness), определяющей вероятность реализации «с точки зрения ультимативных механизмов».
Последние резко разнятся по своего рода «сила притяжения» соответствующих направлений развития к аттрактору «максимума inclusive fitness«, который — и здесь социобиологи снова правы — организации не обойти. Другое дело, что следование этим филогенетическим траекториям — или нет, в отличие от «скорости продвижения», коль последнее уже началось, определяется не ультимативными, а проксимальными механизмами (в чём моё расхождение с социобиологами).
Но даже с их точки зрения не всякая потенциально выигрышная манипуляция будет создана: чтобы отбор «заработал» именно в эту сторону, она должна быть «прибыльней» как минимум текущего состояния «без манипуляции», когда соответствующие поведенческие, морфофизиологические и биохимические феномены существуют просто в виде последствий инициирования и реакции тканей хозяина на жизнедеятельность паразита. Забыв этот важный момент, авторы (как и прочие социобиологи) наивно надеются, что отбор обязательно «усилит», «отшлифует» и «отточит» те последствия паразитирования, что потенциально способны дать манипуляцию. Нет, многие из них так и останутся просто последствиями, именно потому что выработать приспособления отбору «обойдётся дороже», чем не менять ситуацию вообще.
Такие надежды — обычный пример разобранных раньше «ошибок «от логики»: коль есть универсальный механизм формообразования — естественный отбор, и есть возможная точка его приложения, он обязательно включится и «сделает дело», независимо от особенностей меняющейся организации, экологических обстоятельств её изменения и пр. Такая ошибка повсеместна у социобиологов (см. пример с human oestrus). Она создана умонастроением панадаптационизма («парадигма Панглосса»), вообще говоря, несовместным с аналитическим инструментарием данной дисциплины и, увы, часто «выключает» использование последнего — по тем же причинам, по которым позитив от «удачного объяснения» сложной ситуации выключает трезвый скепсис, требующий проверить и взвесить «а оно точно проходит? нет ли другого?». См. «дело о женском оргазме» (его происхождении). Увы, и у Гопко с Михеевым выключило, хотя многие из разобранных ими случаев говорят о необходимости такого трезвения.
Фактически выше Докинз «забыл» о правилах анализа реальности, предложенных своей собственной концепцией, «поддавшись» парадигме Панглосса. А авторы статьи наступили на те же грабли, поддавшись авторитету Докинза. Прим.публикатора]
Манипуляции паразита являются одним из типичных примеров этой концепции, поскольку гены, управляющие проявлением фенотипического признака (например, реакции рачка на свет), находятся в двух разных “машинах выживания” — организме хозяина и организме паразита (Poulin, 2010; Hughes, 2014; Heil, 2016).
Со времени первых работ В. Бетла, Дж. Холмса и Р. Докинза прошло уже несколько десятилетий и за это время паразитические манипуляции превратились в общепринятую и хорошо устоявшуюся научную концепцию (Poulin, 2010). Если поначалу манипуляции поведением хозяина воспринимались как нечто исключительное (Hughes, 2014), то к настоящему моменту принято считать, что они весьма распространены в природе и едва ли не первыми привлекаются в качестве объяснения изменений в поведении хозяина (Parker et al., 2009; Poulin, 2010; Weinersmith, Faulkes, 2014; Moore, 2013).
Адаптивность манипуляций
В 1980-е годы изменения в поведении хозяина было принято рассматривать с точки зрения трех не исключающих друг друга гипотез. Изменения в поведении могли считаться адаптацией паразита — собственно манипуляция, адаптацией хозяина — попыткой избавиться от инфекции и/или ослабить ее негативное влияние на организм и, наконец, просто побочным эффектом инфекции (Moore, 2013). Последнюю гипотезу, кажущуюся на первый взгляд привлекательной, в последнее время все чаще принято отбрасывать, как непродуктивную и не выдерживающую проверку в эксперименте (Moore, 2013). Как ни странно, но при пристальном изучении изменения в поведении зараженного животного, как правило, оказываются адаптацией либо хозяина, либо паразита. Изначальная популярность гипотезы “побочного эффекта инфекции”, вероятно, связана с традиционным биомедицинским взглядом на паразитов как на нечто чуждое, неестественное и вредное. Однако по мере распространения адаптационистских и эволюционистских взглядов на взаимоотношения паразита и хозяина в измененном фенотипе зараженного животного все чаще стали искать и находить адаптации одной из сторон биологического взаимодействия (Мооге, 2013).
[И ведь упорно не видят, что тут нет ничего хорошего, см.выше!]
Согласно современному определению адаптивным считается тот фенотип, который в данных условиях окружающей среды обеспечивает максимальную приспособленность по сравнению с определенным набором альтернативных фенотипов (Lefevre, Thomas, 2008). Другими словами, любые изменения фенотипа хозяина, ведущие к повышению приспособленности паразита, будут поддерживаться отбором и потому являются адаптациями (Lefevre, Thomas, 2008; Poulin, 2010). Действительно, даже если вначале изменения фенотипа хозяина и являются побочным эффектом инфекции, то в дальнейшем естественный отбор будет благоприятствовать тем из них, которые адаптивны, скажем, для паразита, в то время как неадаптивные фенотипические изменения будут элиминироваться (Докинз, 2010, с. 362; Lefevre, Thomas, 2008).
[Напрочь забыта ещё одна категория: адаптивные, но требующие большей «платы», чем текущее состояние побочных эффектов инфекции. А ведь коли оба вида, паразит и хозяин, успешно осуществляют каждый свой жизненный цикл, организация обоих уже адаптивна. И новый отбор, «вырабатывающий манипуляции», включится дополнительно лишь при большей их «прибыльности», «от добра добра он не ищет». Прим.публикатора]
Таким образом, в устоявшейся системе паразит —хозяин, т.е. если паразит и хозяин коэволюционировали достаточно долго, шансы обнаружить побочный эффект инфекции, не подвергавшийся давлению естественного отбора, невелики.
[Вообще, в данном разделе явное противоречие теории авторов, требующей «отбора на уровне генов», и сделанных ими утверждениях о фактах. Неслучайно дальше в статье много разного и интересного о приспособлениях, и совсем ничего — о «генах», которые их хотя бы обуславливают, не говоря уж об «эгоистичных репликаторах» в смысле Докинза, которые их используют «как машину для собственного воспроизводства». Интересно, авторы видят эту невязку с эмпирией у любимой теории? Прим.публикатора.]
Провести четкую грань между фенотипическими изменениями, адаптивными для паразита и хозяина оказывается не так просто по нескольким причинам. Во-первых, не все системы паразит — хозяин удобны для экспериментальной проверки каждого из этих предположений (Moore, 2013). Во-вторых, одно и то же фенотипическое изменение может оказаться одновременно и приспособлением паразита, и приспособлением хозяина. Например, стремление многих насекомых, зараженных паразитами, искать открытые возвышенные и хорошо освещенные солнцем участки может быть объяснено двумя не исключающими друг друга способами (Moore, 2013). С одной стороны, такое поведение делает насекомое более заметной жертвой для следующего хозяина, что выгодно паразиту. С другой стороны, хозяин может оставаться на солнцепеке, чтобы искусственно повысить температуру своего тела и попытаться избавиться от инфекции, поскольку температурная выносливость паразита может быть ниже таковой хозяина (Lefevre, Thomas, 2008; Moore, 2013). Аналогичная смена температурных предпочтений обнаружена также у других видов животных, например, моллюсков (Wesolowska, Wesolowski, 2013) и рыб (Macnab, Barber, 2011). Есть, однако, ряд теоретических рассуждений, свидетельствующий в пользу того, что фенотипические изменения будут гораздо чаще оказываться манипуляциями паразита, а не защитными приспособления хозяина.
Во-первых, жизненный цикл паразита, как правило, короче жизненного цикла хозяина, а численность оставленного им потомства гораздо выше. Таким образом, паразиты способны эволюционировать и вырабатывать адаптивные черты быстрее, чем их хозяева (Sorensen, Minchella, 2001). Второй, более фундаментальный аргумент, связан с применением принципа “жизнь — обед” к эволюционной “гонке вооружений” в системе паразит —хозяин (Dawkins, Krebs, 1979; Докинз, 2010, гл. 4; Lefevre, Thomas, 2008).
[Он неверен, т. к. при прочих равных затраты сторон в эволюционной гонке вооружений сопоставимы, а развивается она за счёт асимметричности «ответов» и «вызовов», при несовершенстве каждой из адаптаций, всегда оставляющей «ахиллесову пяту» в структуре приспособлений. Прим.публикатора]
Под эволюционной гонкой вооружений понимают ситуацию, когда две филогенетические линии (два вида, две четко различимые группы внутри вида и т.п.) коэволюционируют вместе на протяжении долгого времени, причем в ответ на адаптации одной линии другая вырабатывает контрадаптации. Совсем уж строго говоря, в гонке вооружений участвуют два набора коэволюционирующих генов, но здесь мы не будем вдаваться в излишнюю детализацию (Dawkins, Krebs, 1979; Докинз, 2010).
Участниками эволюционной гонки вооружений могут быть паразит — хозяин, хищник — жертва, два конкурента за один и тот же пищевой ресурс, самцы и самки, инвазионные и неинвазионные особи паразитов и т.п. Гонка вооружений может быть симметричной и асимметричной в зависимости от того, одинаково ли направление отбора, действующего на участников (Dawkins, Krebs, 1979; Докинз, 2010). Например, два вида деревьев, конкурирующих в лесу за солнечные лучи, вовлечены в симметричную гонку вооружений, поскольку отбор в этом случае влияет на схожие фенотипические черты у каждого из конкурентов [Нет, в той мере, в какой у них разная структура кроны и мозаика размещения листовых поверхностей в соответствующем объёме, а это именно то чем различаются разные виды деревьев]. В асимметричной гонке вооружений давление отбора влияет на очень несхожие фенотипические черты каждого из участников. Например, давление отбора на хищников, совершенствующее их навыки погони, ловли и убийства, иное, чем давление на жертв, поскольку жертвы совершенствуют в ходе эволюции совершенно иные адаптации: избегание, уклонение, маскировку и т.п.
В гонках вооружений нередка ситуация, когда давление отбора на одну из сторон конфликта сильнее, что позволяет этой стороне развивать более совершенные адаптации и находиться в эволюционной гонке несколько впереди (Dawkins, Krebs, 1979; Докинз, 2010). Принцип “жизнь —обед” как раз иллюстрирует ситуацию, когда давление отбора на одну из сторон в эволюционной гонке вооружений сильнее, чем на другую. Этот принцип получил свое название от изречения Эзопа, написавшего, что заяц бежит быстрее лисицы, поскольку борется за свою жизнь, в то время как она только за свой обед. Другими словами, в некоторых гонках вооружений последствия неэффективно сработавшей адаптации значительно серьезнее для одной стороны, чем для другой. Лисица, не догнавшая зайца, способна после этого оставить потомство и передать свои гены в следующее поколение, в то время, как гены проигравшего в забеге зайца уже никогда не перейдут к его потомкам. Стоит заметить, что принцип “жизнь — обед” вовсе не утверждает, что давление отбора на жертву будет всегда сильнее, чем давление отбора на хищника, как в приведенном выше гипотетическом примере (Докинз, 2010, с. 121—124).
В системе паразит — хозяин давление на паразита, как правило, сильнее, поскольку для паразита-манипулятора попадание в организм следующего хозяина — всегда вопрос жизни и смерти, в то время как для хозяина это не обязательно так (Lefevre, Thomas, 2008). Любой потенциальный хозяин паразита происходит от очень длинной линии предков, многие из которых никогда не встречались с угрозой паразитизма. Более того, многие из организмов-хозяев оставляли потомство до, а иногда и после заражения паразитами. Таким образом, даже у тех генов хозяина, которые не были способны обеспечить эффективную защиту от паразитической угрозы, было немало шансов передаться потомству. Напротив, любой паразит происходит исключительно от тех предков, которые эффективно и успешно решили проблему переноса от одного хозяина к другому (Lefevre, Thomas, 2008). Следовательно, гены хозяина, увеличивающие шансы паразита попасть в организм следующего хозяина, например, гены, отвечающие за манипуляцию, имеют больше шансов закрепиться в генофонде. Хотя в результате эволюционной гонки вооружений “равновесный” фенотип хозяина будет представлять собой нечто промежуточное между оптимальными фенотипами с точки зрения паразита и с точки зрения хозяина, равновесие в системе будет, как правило, смещено в сторону паразитического оптимума из- за большего давления отбора на паразита (Lefevre, Thomas, 2008). Таким образом, обнаружив изменения в поведении зараженного хозяина, мы с большой вероятностью можем утверждать, что столкнулись именно с манипуляцией паразита, а не защитной реакцией хозяина.
Распространенность паразитических манипуляций в природе
Тем не менее вопрос о распространенности паразитических манипуляций в природе остается дискуссионным. Вероятно, не все паразиты являются манипуляторами, однако, (1) какие факторы ограничивают распространение в популяции паразитов генов, отвечающих за манипуляции, и (2) какова доля паразитов-манипуляторов (Poulin, 2010; Ней, 2016)? Среди причин ограничивающих распространение манипуляций наиболее активно обсуждаются затраты на манипуляции. Изменение поведения хозяина, по всей видимости, требует от паразитов немалых затрат вещества и энергии, которые не обязательно окупаются (Poulin, 1994,2010; Brown, 1999; Brown et al., 2003; Poulin et al., 2005; Parker et al., 2009; Heil, 2016 Gopko et al., 2017b. Вопрос о затратах на манипуляции будет обсуждаться в отдельном разделе этой работы.
Хотя число примеров паразитических манипуляций в научной литературе уже исчисляется сотнями, по-прежнему неясно, какова доля видов-манипуляторов от общего числа паразитических видов. Сообщения об отсутствии у каких-то видов паразитов способности к манипуляции относительно редки, но, по всей видимости, это отчасти объясняется нежеланием многих исследователей и научных журналов публиковать отрицательный результат исследования (Poulin, 2010).
[а также вышеизложенной адаптационистской парадигмой, когда всякие изменения после заражения паразитом интерпретируются как «манипуляция», т. к. их выгодность при известной изворотливости ума всегда можно аргументировать, а нуль-гипотеза, что это побочные следствия инфекции, специально не проверяется и не опровергается. Прим.публикатора].
Кроме того, в литературе безусловно встречаются примеры, как “ложноположительных”, так и “ложноотрицательных” примеров манипуляций (Heil, 2016). Ошибочное приписывание паразитам манипуляций может быть связано с недостаточным вниманием к альтернативным объяснениям изменений в поведении хозяина или неаккуратно проведенными экспериментами.
С другой стороны, из того факта, что исследователю не удалось обнаружить манипуляцию в своих экспериментах, вовсе не следует, что паразит, которого он изучал, не является манипулятором. Ученый естественным образом ограничивается лишь небольшим набором поведенческих черт, которые могут меняться под воздействием паразита. Разумеется, нельзя быть уверенным, что для своих манипуляций паразит выбирает именно эти черты. Кроме того, успешная манипуляция вовсе не обязательно должна бросаться в глаза (Heil, 2016). Иногда цель манипуляции сделать зараженного хозяина не отличимым от незараженных особей (Heil, 2016). Например, для паразитов способных горизонтально переноситься от хозяина к хозяину, скажем, возбудителей заболеваний, передающихся половым путем, важно, чтобы зараженная особь не проявляла признаков слабости и не теряла своей привлекательности для потенциальных половых партнеров (Heil, 2016). Это особенно актуально, учитывая, что многие животные активно при спаривании выбирают менее зараженных или более устойчивых к паразитам партнеров (Hamilton, Zuk, 1982; Hamilton et al., 1990). В таком случае, если паразит оказывается способен сделать своего хозяина хотя бы не менее привлекательным для полового партнера по сравнению с незараженной особью, то манипуляцию поведением хозяина можно считать успешной (Heil, 2016). Это вовсе не значит, что зараженное животное не отличается от незараженного. Успех в привлечении полового партнера может быть достигнут путем многократного увеличения производства половых феромонов в организме хозяина по сравнению с незараженными особями (Dass et al., 2011; Vyas, 2013; Heil, 2016), однако в чисто поведенческих тестах такая манипуляция останется незамеченной.
Вообще, паразитические манипуляции, которые не могут быть обнаружены визуально, как правило, выпадают из поля зрения исследователей, и сведения о таких манипуляциях крайне скудны (Moore, 2013). Вероятнее всего это связано с той огромной ролью, которую зрение играет в восприятии человека. Большую часть окружающих нас явлений мы замечаем зрительно, в то время как для паразитов и их хозяев, если это не Homo sapiens, зрение может не играть существенной роли и уступать первенство другим способам воспринимать окружающую действительность. Здесь следует упомянуть такое понятие, как “перцептивный мир” (Umwelt), предложенное Якобом фон Икскюлем (Мооге, 2013). Под ним подразумевается тот окружающий мир, который живое существо способно воспринять с помощью своих органов чувств. Для иллюстрации своей идеи Икскюль использовал эктопаразитических кровососущих клещей. Он заметил, что в поисках пищи клещи реагируют всего на три стимула, причем строго в перечисленном ниже порядке: запах масляной кислоты, температура близкая к 37°С (температуре тела теплокровного животного) и строение волосяного покрова на теле потенциального хозяина (Мооге, 2013). Остальные стимулы не существенны и, вероятно, даже не воспринимаются. Человек и клещ могут находиться в одном и том же уголке леса в один и тот же момент, человек может даже быть для клеща источником пищи, но в то же время их восприятия окружающего мира практически не будут пересекаться (Мооге, 2013).
Для того чтобы полностью исчерпать возможности паразита манипулировать фенотипом своей жертвы и с уверенностью отвергнуть гипотезу манипуляции, необходимо очень глубоко “проникнуть” в перцептивные миры хозяина и паразита. Зачастую такое проникновение приводит к обнаружению новых и неожиданных адаптаций паразита, таких как упомянутое выше использование химических стимулов (половых феромонов) для привлечения хозяина. Еще более яркий пример перцептивного мира, не относящийся напрямую к манипуляциям, способность паразитических тахин (Ormia ochracea) обнаруживать своих жертв — сверчков — по характерному стрекотанию. Необходимость избегать таких паразитов привела к тому, что на нескольких островах Гавайского архипелага, куда вселилась паразитическая тахина, сверчки почти полностью перестали стрекотать буквально за несколько десятков поколений (Zuk et al., 2006). Без проникновения в перцептивный мир паразита исследователям было бы трудно понять, почему на некоторых островах Гавайского архипелага путешественника вместо привычного стрекотания встречает почти полная тишина.

Cверчок Teleogryllus oceanicus
По всей видимости, именно исследование манипуляций, затрагивающих фенотипические черты хозяев, не заметные для человеческого глаза, является одним из наиболее интересных направлений дальнейших исследований. Впрочем, далеко не единственным.
В завершение отметим, что, хотя вопрос о распространенности паразитических манипуляций далек от окончательного решения, в настоящий момент принято считать, что встречаются как паразиты-манипуляторы, так и виды (и даже группы в рамках одного и того же вида) не манипулирующие поведением хозяина (Poulin, 2010). При этом несомненно, что манипуляции являются весьма распространенным природным феноменом и число манипуляций, не попавших в поле зрения исследователей, вероятно, весьма велико (Poulin, 2010; Мооге, 2013; Ней, 2016).
Несостоявшийся “кризис жанра” и новые направления исследований
Необходимо сказать несколько слов о том, в каком направлении развивается изучение паразитических манипуляций в последние годы и какую роль играет манипуляторная тематика в современной эволюционной, поведенческой и экологической науке. Любопытно, что почти два десятилетия назад известный паразитолог Пулен высказал предположение (Poulin, 2000), что интерес к концепции паразитических манипуляций начинает “выдыхаться”. Проведенный им мета-анализ показал, что с начала 1970-х годов, когда паразитические манипуляции впервые попали в поле зрения ученых, по конец 1990-х, статистический размер эффекта (effect size) в работах, посвященных манипуляциям, имел тенденцию к снижению. Объяснить это явление можно было как исчерпанием самого предмета исследований, так и потерей интереса научного сообщества к этой теме, т.е. выходом ее из моды (Poulin, 2000).
Однако 16 лет, прошедших со времени выхода этой работы, характеризовались бурным всплеском интереса к паразитическим манипуляциям фенотипом хозяина. Были обнаружены новые впечатляющие примеры этого феномена (см. Seppala et al., 2004, 2005; Yanoviak et al., 2008; Mikheev et al., 2010; Maure et al., 2011, 2013 и пр.). Намечены совершенно новые направления исследований, например манипуляции паразитов фенотипом растений (Ней, 2016), манипуляции неинвазионных паразитов (Parker et al., 2009; Dianne et al., 2011; Hafer, Milinski, 2015; Gopko et al., 2015), конфликт и кооперация паразитов-манипуляторов в организме хозяина (Thomas etal., 2005; Hafer, Milinski, 2016; Weinersmith et al., 2014; Gopko et al., 2017b). Паразитические манипуляции все больше интересуют матбиологов и мета-аналитиков, пытающихся предсказать условия, при которых манипуляция может быть поддержана естественным отбором, каких стратегий манипулирования придерживаются паразиты на разных этапах своего жизненного цикла, как физиологические механизмы контроля над поведением хозяина отличаются у паразитов-генералистов и паразитов-специалистов (Fenton, Rands, 2006; Parker et al., 2009; Vickery, Poulin, 2010; Fredensborg, 2014). Наконец, только в последние несколько лет экологи оценили, какую важную роль паразиты в целом и паразитические манипуляции в частности могут играть в различных экосистемах, сближая трофические уровни, ускоряя перенос вещества и энергии по трофическим цепям, а подчас и являясь важным пищевым ресурсом для других живых существ (см. разд. IV этого обзора).
II. Основные стратегии манипулирования. “Классическая манипуляция” (манипуляция неинвазионных паразитов)
Жизненные циклы паразитов разнообразны, и столь же разнообразны могут быть их стратегии манипулирования. В этом разделе мы кратко опишем, какие цели может паразит преследовать своей манипуляцией в зависимости от особенностей его жизненного цикла, и подробно остановимся на манипуляциях паразитов, передающихся по трофической цепи.
Паразитов можно разделить на две категории. Одни способны провести всю жизнь на одном хозяине или на одном виде-хозяине (моноксенные паразиты), т.е. в относительно постоянных условиях. Другим для завершения жизненного цикла необходима смена видов-хозяев (гетероксенные паразиты) или же они часть жизни проводят, паразитируя на других организмах, а другую часть — как свободноживущие организмы (паразитоиды). Гетероксенные паразиты и многие паразитоиды резко меняют условия обитания в ходе своего жизненного цикла. Стратегии манипулирования этих категорий паразитов могут существенно различаться (Poulin, 2010; Михеев, 2011).
Манипуляции моноксенных паразитов
Хотя моноксенный паразит может провести всю жизнь на одном индивидууме, это вовсе не означает, что ему за время своей жизни не нужно сталкиваться с другими представителями вида-хозяина. Такая необходимость может возникнуть, если паразит или его потомство передается следующему хозяину при непосредственном контакте, например половым путем, при прикосновении или укусе. Теоретически такие паразиты заинтересованы в большей частоте контактов между особями вида-хозяина, т.е. манипулируют зараженным хозяином так, чтобы он как можно активнее искал контактов с другими особями своего вида (Dawkins, 1999; Lefevre, Thomas, 2008; Poulin, 2010; Михеев, 2011). Несмотря на логичность подобного предположения, примеров таких манипуляций обнаружено немного, вероятно, из-за недостаточной изученности вопроса (Poulin, 2010; Heil, 2016). Известна, однако, способность передающихся половым путем паразитов увеличивать сексуальную привлекательность своего хозяина (см. подробный обзор примеров в Heil, 2016). Например, такой распространенный, в том числе и в человеческой популяции, паразит, как Toxoplasma gondii (Flegr et al., 2014a, b) увеличивает выработку половых гормонов у самцов крыс, делая их более сексуально привлекательными для самок (Dass et al., 2011). При этом известно, что у крыс, кроликов, овец, собак, а возможно, и у людей токсоплазмоз способен передаваться половым путем (Flegr et al., 2014а, Heil, 2016). Тем самым увеличение частоты половых контактов носителя инфекции повышает и шансы паразита заразить нового хозяина.
Токсоплазма Гонди
Известный пример манипуляций, связанных с половым поведением, обнаружен в системе паразитический гриб (Entomophthora muscae) — комнатная муха (Musca domestica) (Moller, 1993). Споры Е. muscae прорастают гифами в абдомене мухи, приводя к смерти хозяина. Мухи часто умирают на открытых местах, оказываясь хорошо заметными для других представителей своего вида. Даже мертвые они по какой-то причине сексуально привлекательны для других особей того же вида, в первую очередь самцов (Moller, 1993). Несмотря на то что погибшая от инфекции муха окружена пучками конидий, она является источником каких-то зрительных или химических стимулов, которые заставляют других мух приближаться к ней, касаться ее и в конечном итоге с очень большой вероятностью (> 90%) заражаться спорами гриба-энтомофага (Moller, 1993). Еще один патоген, предположительно манипулирующий поведением хозяина с целью увеличить частоту его контактов с другими особями своего вида, возбудитель бешенства Rabies lyssavirus. Именно с манипуляциями иногда связывают повышенную активность и агрессивность животного (Dawkins, 1999; Lefevre, Thomas, 2008; Poulin, 2010). Впрочем, повышенная агрессивность далеко не единственная форма проявления бешенства и для того, чтобы подтвердить гипотезу манипуляции поведением зараженного животного требуются дополнительные исследования (Poulin, 2010).

Entomophthora muscae, колонизировавшая навозную муху Scathophaga stercoraria
В некоторых случаях моноксенным паразитам необходимо покидать своего хозяина для поиска полового партнера или откладки яиц. Так, поступают например эктопаразитические рачки р. Argulus, обитающие на коже рыб. Хотя аргулюсы облигатные паразиты и во взрослом состоянии питаются исключительно тканями и кровью хозяина (Mikheev et al., 2000; Walker et al., 2011), они всю жизнь сохраняют способность к активному плаванию и часто меняют хозяев (Mikheev et al., 2015). Логично предположить, что паразиту также выгодно, чтобы зараженная им рыба держалась ближе к другим рыбам в стае. Показано (Михеев, 2011), что в стаях радужных форелей (микижей), зараженных Argulus foliaceus, рыбы предпочитали держаться ближе друг другу по сравнению с контрольными стаями, состоящими из незараженных рыб. Кроме того, рыбы, зараженные аргулюсом, значительно снижали скорость плавания и становились менее агрессивными по сравнению с контрольными рыбами (Михеев, 2011). Вероятнее всего, эти изменения в социальном поведении микижи вызваны выделением в воду веществ тревоги из клеток, поврежденных рачком при питании (Mikheev et al., 2015). Таким образом, в этом случае паразит использует поведенческие механизмы, обычно служащие рыбам для защиты от хищников, чтобы изменить групповое поведение рыб к собственной выгоде.

Сосательный аппарат и присоски, с помощью которых рачок-карпоед Argulus foliaceus прикрепляется к телу рыб. Фото с сайта http://www.aquarienfotografie.net
Наконец, в тех случаях, когда моноксенный паразит не делает ставку на непосредственный контакт между хозяевами для передачи инфекции, он может тем не менее менять место его обитания (habitat niche), чтобы увеличить шансы заразить других хозяев. В качестве примера можно выбрать уже упомянутый ниже паразитический гриб Е. muscae и еще одного его хозяина — навозную муху (Scathophaga stercoraria). Зараженные мухи перед смертью занимают положение, наиболее благоприятное для распространения спор гриба (Maitland, 1994). Они прочно прикрепляются к нижней стороне листа близко к его кончику. При этом местом своего упокоения зараженные мухи, как правило, выбирают относительно большие, возвышающиеся над другими травянистые растения, умирая поблизости от их верхушки. Наконец, они соотносят место, где замирают перед смертью, с розой ветров, располагаясь преимущественно с подветренной стороны побега (Maitland, 1994).
Для сравнения: здоровые мухи предпочитают ползать по верхней поверхности листа, в основном обитают на низкорослых травянистых растениях, а вокруг растений и на них они распределены случайным образом, т.е. направление ветра для них несущественно (Maitland, 1994). Предполагается, что паразит манипулирует предсмертным поведением своего хозяина, чтобы после смерти он оказался максимально эффективным распространителем спор гриба. Кроме того, расположение на нижней стороне листа, вероятно, способствует созданию оптимальных условий для развития спор (локальное повышение влажности) и защищает паразита от смывания на землю дождем (Maitland, 1994).
Пожалуй наиболее известным примером моноксенного паразитического гриба-манипулятора служит Ophiocordyceps unilateralis — группа видов, поражающая различных тропических муравьев. Вызванные этим патогеном поведенческие изменения настолько впечатляющи, что зараженные О. unilateralis муравьи получили в научной литературе название — “муравьи-зомби” (zombie-ants) (Hughes et al., 2011b; Evans et al., 2011; Weinersmith, Faulkes, 2014). Заражение начинается с того, что спора гриба попадает на поверхность экзоскелета муравья и проникает внутрь тела хозяина, сочетая механическое давление и химическое (смесь ферментов: хитиназ, липаз и протеаз) воздействие на его покровы и успешно избегая его защитных механизмов (Evans et al., 2011). В дальнейшем дрожжеподобные клетки распространяются по телу хозяина и прорастают гифами, особенно сильно разрастаясь в головной капсуле муравья, что играет ключевую роль в успехе последующей манипуляции его поведением (Hughes et al., 2011b). На этом этапе инфекция протекает бессимптомно.

Рис. 1. Муравьи-древоточцы, убитые паразитическим грибом Ophiocordyceps unilateralis в природе (на изображениях A и B видны грибы, выросшие из затылков жертв) и в лаборатории (D, E). A, D — муравьи Camponotus castaneus, B, E — C. americanus. C — культура паразитического гриба, полученная из зараженной особи C. castaneus. Изображение из обсуждаемой статьи в BMC Evolutionary Biology
Спустя несколько дней паразит начинает выделять в гемоцель муравья вещества, которые заметно меняют его поведение (Evans et al., 2011). Муравей покидает свою колонию в кроне дерева и спускается во влажный подлесок, чего здоровые муравьи обычно не делают. Любопытно, что поведенческие изменения у всех муравьев синхронизированы и приурочены к довольно короткому промежутку времени (двум предполуденным часам и полудню), словно паразит включает в организме хозяина будильник (Hughes et al., 2011b). Оказавшись в подлеске, зараженный муравей начинает мучительное путешествие в поисках места, наиболее подходящего для дальнейшего развития паразита. Несмотря на периодические спазмы, он может миновать несколько невысоких растений и “исследовать” несколько десятков и даже сотен (до 239) листьев в поисках подходящего (Hughes et al., 2011b)
Определившись с выбором, он забирается на абаксиальную, т.е. внешнюю по отношению к центральной оси побега сторону листа, и впивается в его центральную жилку (Andersen et al., 2009; Hughes et al., 2011b). Затем мышечные ткани челюстей муравья быстро дегенерируют, причем за процесс атрофии, видимо, ответственны гифы гриба, обильно обволакивающие мозг и мышечные волокна хозяина. После этого муравей, хотя и остается в живых еще несколько часов, уже не способен разомкнуть челюсти, хватка которых становится мертвой (Hughes et al., 201 lb). После смерти муравья гифы гриба полностью опутывают тело хозяина, мумифицируя его и прочно прикрепляя к субстрату, а спустя несколько недель плодовое тело О. unilateralis прорастает через его головную капсулу, тем самым завершая жизненный цикл паразита (Evans et al., 2011). Остается заметить, что “мертвая хватка” зараженного муравья настолько сильна, что преодолевает геологические эпохи. Характерные отверстия, оставленные на листе мандибулами муравья, были обнаружены в окаменелости возрастом почти 50 млн лет (Hughes et al., 2011а).
В завершение отметим, что, несмотря на эффектность приведенных выше примеров, манипуляции моноксенных паразитов изучены в целом меньше, чем гетероксенных. Например, количество убедительных примеров манипуляций паразитов, передающихся при непосредственном контакте (укус, касание, половой акт), до сих пор невелико. То же справедливо и для паразитов, передающихся половым путем (Poulin, 2010; Ней, 2016). Неясно, свидетельствует ли обнаруженный паттерн о недостаточной изученности вопроса или же имеет под собой более глубокие эволюционные основания. Например, давление отбора на паразитов, передающихся при непосредственном контакте, может быть не столь велико: вероятность передачи инфекции достаточно высока и при обычном, не измененном паразитом поведении хозяина (Poulin, 2010).
Манипуляции паразитоидов и гетероксенных паразитов
Стратегии манипулирования паразитоидов и гетероксенных паразитов разумно рассматривать совместно (Poulin, 2010), поскольку жизненный цикл этих организмов характеризуется одной важной общей чертой. И те, и другие в ходе своего жизненного цикла резко меняют условия своего существования: паразитоиды переходят от жизни в организме хозяина к свободному существованию во внешней среде, а гетероксенные паразиты меняют хозяев, нередко заметно различающихся как по особенностям физиологии, так и по месту своего обитания. Например, жизненный цикл большинства видов трематод включает трех хозяев (Galaktionov, Dobrovolskij, 2003). Так, для D. pseudospathaceum и Euhaplorchis califomiensis — двух известных трематод-манипуляторов — первыми промежуточными хозяевами служат брюхоногие моллюски, вторыми — рыбы, а окончательными — рыбоядные птицы (Seppala et al., 2004; Lafferty, Morris, 1996). Другими словами, чтобы успешно завершить свой жизненный цикл, этим трематодам нужно побывать в организмах животных, относящимся к двум разным типам, и сменить среду своего обитания с водной на воздушную. Для решения этой задачи паразитам требуется большой набор адаптаций, к числу которых относятся и изменение фенотипа хозяина.
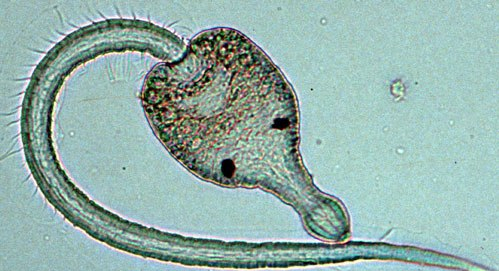
…и сама трематода (источник тот же)
Вероятно, именно поэтому среди паразитоидов и гетероксенных паразитов обнаружено большинство видов-манипуляторов (Poulin, 2010). Пулен (Poulin, 2010) предложил выделить несколько стратегий, в которые хорошо укладывается большинство примеров манипулирования, обнаруженных к настоящему моменту. Ниже его классификация дается с некоторыми изменениями — например, мы отдельно описали манипуляции моноксенных паразитов (см. выше) и не будем к ним возвращаться.
Первую стратегию применяют паразитоиды, резко меняющие условия обитания в ходе своего жизненного цикла. Как правило, этот переход сопряжен со смертью хозяина, причем, что характерно, время и/или место смерти обычно точно определяется паразитом (Maure et al., 2011). В этом смысле манипуляции паразитоидов во многом напоминают описанные выше манипуляции моноксенных паразитов. Цель манипуляции многих паразитоидов — заставить хозяина кардинально изменить место своего обитания зачастую на совершенно неподходящее. Хорошим примером служат волосатики (Nematomorpha, класс Gordoidea), личинки которых обитают в теле насекомых, в то время как взрослые стадии этих червей — свободноплавающие животные. Взрослые особи волосатика выметывают яйца в воду, где в течение 2—4 нед из них развиваются личинки. Личинки не способны к активному плаванию и могут быть проглочены различными водными животными. Многие из этих животных оказываются неподходящими хозяевами для паразита — тупиковыми хищниками (deadend predators (Seppala et al., 2008)) (Hanelt et al., 2016, nematomorpha.net). Другие организмы, например личинки водных насекомых, служат волосатикам дополнительными хозяевами (Biron et al., 2005 а; Thomas et al., 2002).
Оказавшись в организме такого хозяина черви инцистируются и в таком состоянии способны пережидать продолжительное время. Со временем личинка превращается во взрослое насекомое и оказывается на суше, где рано или поздно погибает, а ее тело может быть съедено окончательным хозяином. Так, сверчки Nemobius sylvestris, до трети рациона которых составляют мертвые насекомые, заглатывают цисты волосатика Paragordius tricuspidatus с пищей, и в их организме паразит развивается во взрослое животное — тонкого длинного червя, занимающего почти всю полость тела хозяина (Thomas et al., 2002; Biron et al., 2005a). Чтобы завершить свой жизненный цикл, волосатику необходимо вернуться в водоем. Очевидная сложность заключается в том, что сверчки — наземные животные и редко по своей воле оказываются в водоеме. Здесь и возникает необходимость манипулировать поведением хозяина, чтобы заставить его очутиться в воде. Тома с соавт. (Thomas et al., 2002) экспериментально показали, что зараженные волосатиком сверчки, помещенные рядом с наполненной водой кюветой, оказывались в воде достоверно чаще, чем незараженные сверчки. По всей видимости, волосатики химически воздействуют на нервную систему хозяина (показано на примере кузнечика Месопета thalassinum), заставляя его искать воду и в конечном итоге совершать акт суицида, бросаясь в водоем (Biron et al., 2005b).
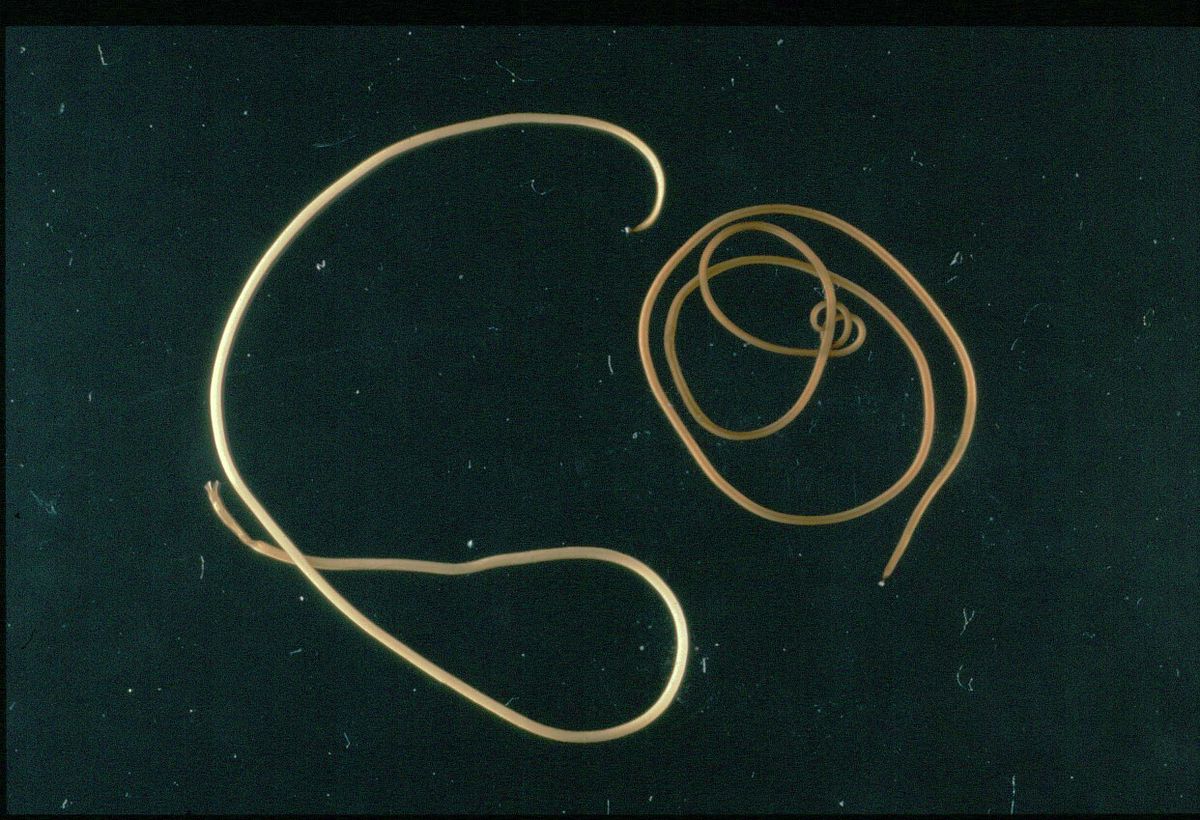
Волосатик Paragordius tricuspidatus
Другой пример — паразитические перепончатокрылые — наездники. Здесь можно привести множество примеров манипуляций фенотипом хозяина, но мы ограничимся лишь двумя. Так, костариканский наездник Hymenoepimecis argyraphaga, относящийся к семейству ихневмонид, паразитирует на пауках Plesiometa argyra. Обычно эти пауки плетут большую ажурную паутину, в центре которой и подстерегают добычу (Eberhard, 2001). Наездник атакует паука, временно (на 10—15 мин) парализует его и откладывает яйцо на поверхность его брюшка (абдомена) (Eberhard, 2001). Яйцо плотно приклеивается к абдомену и вскоре из него выходит личинка наездника. Личинка проделывает несколько небольших отверстий в брюшке паука, через которые и питается гемолимфой хозяина. Кроме того, она, видимо, впрыскивает в организм паука какие-то химические вещества, которые в конечном итоге меняют его поведение. Однако в первые 7—14 дней после заражения эти изменения незаметны, и лишь в последнюю ночь перед окукливанием личинки наездника паук начинает вести себя иначе (Eberhard, 2000, 2001). Он начинает строить новую паутину, однако она совсем не похожа на ту ажурную ловчую сеть, что он строил раньше. Вместо этого он строит небольшой, но прочный каркас из нескольких толстых нитей и хорошо укрепляет его центр (Eberhard, 2000, 2001).

Найдите десять отличий! Слева паутина, сплетенная «обычным» пауком Plesiometa argyra, а справа его зараженным паразитом собратом. На правом фото хорошо заметен, свисающий из центра паутины кокон. Источник фото – Eberhard, 2001 (http://slideplayer.com/slide/6625943)
Когда работа паука завершена, личинка наездника убивает и поедает его. Затем она забирается в центр паутины, где плетет кокон, в котором окукливается, а еще через неделю из кокона выходит имаго наездника. Вероятно, цель манипуляции поведением хозяина защитить паразита от сильных тропических ливней, способных повредить кокон, в котором наездник проходит метаморфоз (Eberhard, 2000, 2001). Любопытно, что если в последнюю ночь перед смертью хозяина паразита удалить с абдомена паука, то хозяин выживает, но продолжает плести паутину неправильной формы еще в течение недели (Eberhard, 2001). Пo всей видимости, все это время паук находится под действием каких-то химических веществ, выделенных паразитом в его гемолимфу (Eberhard, 2001). Стоит отметить и то, как именно паразит меняет поведение хозяина. Он не заставляет паука делать нечто новое. Вместо этого паук совершает движения, очень похожие на плетение нормальной паутины, однако он не переходит от одного типа движений к следующему. Вместо этого он раз за разом повторяет одни и те же движения, что и приводит к появлению толстых, сотканных из множества тонких, наложенных одна на другую нитей. Другими словами, манипуляция заключается не в том, что паразит вызывает у хозяина экспрессию новых поведенческих черт, а в том, что он подавляет часть старых, необходимых для плетения нормальной паутины (Eberhard, 2000, 2001).
Другой пример — Dinocampus coccinellae, наездник, паразитирующий на божьих коровках, в том числе на особях вида Coleomegilla maculata. Наездник откладывает в гемоцель своей жертвы яйцо, из которого в течение примерно двадцати дней развивается личинка, готовая покинуть тело хозяина (Maure et al., 2011, 2013; Dheilly et al., 2015). В течение этого времени поведение божьей коровки остается обычным. Однако к моменту, когда личинка покидает организм хозяина, поведение хозяина меняется. Божья коровка становится малоактивной, почти неподвижной, но периодически все ее тело подрагивает (тремор). При этом личинка наездника окукливается между конечностями хозяина. Таким образом, божья коровка словно прикрывает своим телом куколку паразита. Спустя неделю, наездник завершает метаморфоз и улетает (Maure et al., 2011, 2013). Любопытно, что хозяин все это время остается живым, и теоретически может восстановиться после заражения паразитом и его манипуляции. В условиях лаборатории выживало до четверти божьих коровок, но трудно оценить, каковы шансы хозяина восстановиться после инфекции в естественных условиях (Maure et al., 2011). Такая невысокая вирулентность паразита для божьих коровок, возможно, связана с необычным механизмом манипуляции паразита.
Откладывая яйцо в божью коровку, наездник одновременно заражает ее вирусом (DcPV), обитающим в яйцеводе насекомого. Дальнейшие изменения в поведении хозяина связаны с воздействием на его организм вируса и реакцией иммунитета на этот вирус (Dheilly et al., 2015). Отметим, что опосредованное воздействие паразита на нервную систему хозяина через его иммунитет считается одним из способов манипулирования паразита поведением хозяина (Lafferty, Shaw, 2013; Cezilly et al., 2014), хотя говорить об этом с уверенностью трудно, поскольку в настоящий момент механизм манипуляций известен лишь для относительно небольшого числа систем паразит — хозяин.
Оба приведенных примера характерны тем, что паразит не просто использует ресурсы хозяина для собственного роста и развития, но и заставляет его выполнять функции телохранителя (bodyguard manipulations) (Maure et al., 2011, 2013). В первом случае хозяин защищает паразита косвенно, сплетая жесткий каркас, на который личинка наездника подвешивает свой кокон. Во втором случае божья коровка защищает паразита непосредственно своим телом. В обоих случаях цель манипуляции поведением хозяина — это защита наиболее уязвимых жизненных стадий паразита. Подобные защитные манипуляции относительно недавно привлекли внимание ученых. Однако есть основания полагать, что в природе они нередки (Parker et al., 2009). Более подробно они будут рассмотрены в разд. III (см. Манипуляции неинвазионных паразитов).
Вторая стратегия манипулирования применяется паразитами, использующими хозяина для производства и распространения своих расселительных стадий. Типичный пример — трематоды, промежуточным хозяином которым за редкими исключениями служит моллюск (Galaktionov, Dobrovolskij, 2003). В моллюска проникает покрытая ресничками личинка ми рацидий, которая превращается в материнскую спороцисту. Материнская спороциста дает начало либо дочерним спороцистам, либо редиям, которые способны путем апомиктического партеногенеза (Галактионов, 2016) производить мириады (до десятков тысяч в сутки) своих генетических клонов — церкарий (Lyholt, Buhmann, 1996; Taskinen, 1998; Soldanova et al., 2016). Для паразита это долгосрочный проект, поскольку моллюск может годами служить инкубатором для потомства паразита (Karvonen et al., 2006). Манипулируя фенотипом хозяина, такая трематода преследует несколько целей: во-первых, она старается максимизировать производство своих расселительных стадий, во-вторых, по мере возможности пытается уменьшить вероятность гибели хозяина в результате неблагопр
Свежие комментарии