 Print
Print  PDF
PDF
Самый крупный из ныне живущих дятлов Mulleripicus pulverulentus (после исчезновения белоклювого Campephilus principalis), тоже относится к коммунально гнездящимся. Здесь этот образ жизни развился из кормления в стаях, часто многовидовых, вообще присущего тропическим дятлам
Одиночный — не значит «несоциальный», и наоборот
Содержание
Благодаря Лене Клещенко почитал «Сэкономить на мозгах», что у дятлов «социальные» виды имеют относительно меньший размер мозга, чем «парные», те — меньше, чем одиночные, что резко контрастирует с общей тенденцией, показанной у млекопитающих и птиц. И мне тут за дятлов обидно стало..
Тут я засомневался — не столько в «дятлах», хотя выборка видов, просмотренных авторами исходной статьи Fedorova et al., 2015 , пусть многочисленна, но неполна с т.з. выделенных ими категорий (см. дополнительные материалы к ней). Да, «социальные» виды — т. е. для авторов с коммунальным гнездованием и помощниками, живущие в группах более обширных чем особи с их потомством — выбраны почти все. Хотя социальная жизнь тропических видов, которых в этом семействе большинство, до сих пор исследована неважно, примерно у половины совсем неизвестна, а как эпизод помощничество встречается и у наших обычных видов вроде малого дятла Dendrocopos minor (увы, отнесённого ими к «одиночным»).
И социальный полиморфизм они полностью игнорируют, а ведь именно у дятлов он полностью переворачивает их классификацию. Групповое гнездование с помощничеством оказывается крайним случаем «одиночного», реализующемся при больших и устойчивых запасах корма, с высокой плотностью, не позволяющей молодым немедля занять пустующие «вакансии» гнездовых территорий и обрекающие остаться в группах с «ожиданием» подъёма по тамошней иерархии с превращением из помощника в размножающуюся птицу. И весь этот континуум видим у самого знаменитого из «социальных» дятлов — желудёвого Melanepres formicivorus .
Действительно, в группах этого и других «социальных» видов дятлов самцы и самки ведут себя предельно атомизированно и «эгоистично» в главном, что свело их в «коммуну» — в размножении.
Если территорию они защищают вполне кооперативно, то корм собирают независимо друг от друга, кормят птенцов уже предельно конкурентно — вплоть до того что одновременная встреча у гнезда часто завершается дракой и/или съеданием корма перед ней вместо того чтобы отдать птенцам. Со снесением яиц ещё хуже — делающие это позже склонны уничтожать яйца напарниц, вместо ожидаемого «разделения труда» по насиживанию (тем более что если она их отстоит, насиживают всё одно вместе).
То же самое видим в парах у «одиночных» видов, вроде больших пёстрых дятлов Dendrocopos major, где после вылета птенцов пары распадаются уже в период вождения выводка. Общая территория пары делится на одиночные участки самца и самки даже в случае, если родители осенью и зимой держатся в части прежнего, более обширного участка, а не перемещаются в конце лета — осенью в другие биотопы, более пригодные для зимовки. Первое у D.major обычно в зоне широколиственных лесов, при круглогодичном питании насекомыми, второе — северней, в смешанных и бореальных лесах, где с августа по апрель они питаются семенами хвойных, что требует регулярных и достаточно дальних перемещений дважды в год из зимовочных биотопов в гнездовые и обратно. См.видовой очерк в «Птицах России…».
А вот виды с многолетне-постоянными парами и персонализированными связями партнёров, поддерживаемыми круглогодичным общением с «перебиранием пёрышек» и вне сезона размножения (без него пара распадается, после чего теряет и территорию) у белоспинного Dendrocopos leucotos, трёхпалого Picoides tridactylus, чёрноспинного трёхпалого P.arcticus, малого, пушистопёрого D.pubescens, желны Dryocopus martius — ведут себя совершенно иначе. Они как будто «замыкаются друг на друге»: соседей и/или пришлых «третьих» особей пара в основном отгоняет, независимо от характера их «претензий» — отхватить часть территории пары, как у пушистопёрого, или спариться с самкой/самцом, как в трио трёхпалого/малого. Подобные трио появляются регулярно, но неизменно временны (несколько дней-неделя), это флюктуации типа воронки в ванне, устойчивых социальных объединений на них виды с постоянными парами не строят. См. Фридман, Ерёмкин, 2003 про взаимоотношения с «дополнительным» самцом, и Dorka, 1996 — с самкой.
Все эти подробности отлично известны специалистам по группе, давно описаны и пр. Увы! Авторы статьи к ним не относятся, а знающих не спросили, почему все вышеперечисленные виды в их классификации — не «парные», а «одиночные». Это при том, что реакция пары пушистопёрых дятлов на нарушителей как единого целого, с исследованием обмена акустическими сигналами, поддерживающего согласованную выгонку нарушителя, активно исследуется с 60-х гг., и последняя статья про это вышла только что (Schuppe et al., 2016).
Но главное, неверна сама идея формальной классификации «социальных/несоциальных» видов, когда живущие в группах — «социальные», в парах — «парные», а вроде бы вдали друг от друга на достаточно обширных территориях — «одиночные». Сегодня уже понятно, что «рядом» — не значит «вместе» и «солидарно»; вид может устойчиво жить скученно и быть несоциальным, страдать от социальной дезорганизации, как «колониальные» полуденные песчанки или те же желудёвые дятлы в брачном аспекте «общественной жизни» (но не в территориальности»).
«Социальность» как -связанность и -зависимость индивидов, пусть «одиночных»
И наоборот, вроде бы одиночные дневные песчанки Psammomys obesus не менее социальны, чем плотно живущие и строящие подземные «города» большие песчанки Rhombomys opimus (как выясняется, близкородственные первым). Или вроде бы «одиночные» кошачьи не менее социальны, чем живущие в группах псовые, см. М.Н.Ерофеева, С.В.Найденко. «Пространственная организация популяций кошачьих и особенности их репродуктивных стратегий«. Просто мнимо одиночные виды общаются и регулируют поведение друг друга (т.ч. разные особи ведут себя не независимо, а кооперативно — главный, как дальше увидим, признак «социальности»), больше при помощи дистантных и долговременно действующих сигналов — запахов, других меток, формирующих сигнальное поле, нежели непосредственными контактами с обменом позами/вокализацией. Хотя если численность или экологические обстоятельства заставляют скапливаться, кошки не меньше способны устанавливать отношения, дифференцировать статусы и пр.
Нет, «социальность» vs «несоциальность» не то же самое, что «групповой» vs «одиночный» образы жизни, и в общем случае это даже вполне независимые категории, а не пересекающиеся. Те же асоциальные полуденные песчанки Meriones meridianus могут жить очень скученно из-за обилия корма на сенобазах, почему их (неверно) назвали «колониальными). Однако зверьки здесь дифференцируют время и место активности так, чтобы минимизировать встречи; если контакты случаются, в них почти нет ритуализованного поведения, только некоторый интерес, потом страх или злоба, потом агрессия, победа и поражение в которой «не имеет последствий». Победитель и побеждённый уже в следующей встрече ведут себя так как будто ничего не было, случившееся не меняет долговременно их активность — ни по составу, ни по характеру притязаний.
У социальных видов, тех же больших и монгольских песчанок, наоборот. Они активно ищут контактов, в которых преобладают ритуализованные формы поведения с существенной долей неагрессивных моментов. Выигрыш и проигрыш имеют существенное последействие, надолго формуют поведение и победителя и побеждённого. Чем «социальнее» вид, тем больше и дольше последствия, см. подробнее «Грызуны в рукопашном бою» и «Синдром социальности…«. Да и в территориальных конфликтах дятлов фиксируется то же самое.
А благодаря сигнальному полю группировки прошедшие взаимодействия меняют поведение и третьих особей — «зрителей», почему каждый контакт выливается во взаимодействие «всех со всеми». У больших пёстрых дятлов и других вроде бы одиночно-территориальных видов это происходит на многажды большей площади поселений, чем у мелких млекопитающих, почему эти виды считаются «одиночными», хотя предельно социально связаны и социально зависимы в рамках целого, существующего — важный момент! — независимо от персональных смен в составе сообщества. см. «Победы и поражения большого пёстрого дятла» и «Зачем большим пёстрым дятлам охранять территории?»

Демонстрации территориальной агрессии больших пёстрых дятлов
Как показывает А.В.Чабовский в «Эволюции социальности грызунов открытых пространств: экологические аспекты» (2006) действуют, обеспечивая «связанность» и «зависимость», нужные для социальности, не особи друг на друга, а социальная среда группировки — на всех. В т.ч. чужаков, активных вселенцев, пробующих занять территорию/образовать пару/получить статус в данной группировке). Социальность — вернусь снова к работе Чабовского — определяется «не числом, а умением»: плотностью и интенсивностью контактов, обеспечивающей социальную связанность индивидов, входящих в социум, и согласованность их поведения между собой. В меру последней участки разных зверьков (вроде бы «одиночных») оказываются скоординированными на местности, совпадающими или взаимодополнительными, не случайно разбросанными друг относительно друга.
 И поведение в социальных контактах, даже случившихся относительно случайно, оказывается не независимым, заданным только их собственными хотелками, в смысле «внутренним» возбуждением каждой особи, её собственными агрессией и сексуальности, но той дифференциацией поведенческих ролей, которая уже случилась в прошлых контактах (скажем, разделение на доминанта и подчинённого, или резидента, отстоявшего участок, и проигравшего на его границах захватчика), устойчиво держится посейчас, и корректировка которой возможна лишь через инициирование новых контактов, и их выигрывание прежними проигравшими и наоборот.
И поведение в социальных контактах, даже случившихся относительно случайно, оказывается не независимым, заданным только их собственными хотелками, в смысле «внутренним» возбуждением каждой особи, её собственными агрессией и сексуальности, но той дифференциацией поведенческих ролей, которая уже случилась в прошлых контактах (скажем, разделение на доминанта и подчинённого, или резидента, отстоявшего участок, и проигравшего на его границах захватчика), устойчиво держится посейчас, и корректировка которой возможна лишь через инициирование новых контактов, и их выигрывание прежними проигравшими и наоборот.
Сама по себе социальная асимметрия, раз установившись между исходно «равными» или «одинаковыми» особями, «не пройдёт», не забудется, даже если одну из них отловить и держать в неволе, так что её участок займут вселенцы или разделят соседи (правда, недолго, не больше нескольких дней; я так в 80-х гг. делал с большими пёстрыми дятлами, содержа их в вольере на балконе). «Третьи» особи не дадут «стереть» уже случившуюся дифференциацию, благодаря которой формы поведения у всех особей в сообществе больше согласуются с их ролью/статусом — а значит, теми особенностями проблемных ситуаций, которые данная особь создаёт для других компаньонов — чем с их внутренним состоянием или экологическими обстоятельствами.
Чтобы эти взаимосвязанность и взаимозависимость между «развёртками» поведения индивидов во времени и в пространстве а) были и б) поддерживались устойчиво на всей территории сообщества несмотря на разнообразные «помехи» извне и «напряжения» изнутри, отнюдь недостаточно единичных взаимодействий между ними, даже самых результативных.
Их специфический результат — то самое разделение на выигравших и проигравших в каждом взаимодействии каждого контекста: ухаживания, охраны территории, агрессивного доминирования и пр. должны кумулироваться, «распространяться» по социуму через ретрансляцию «зрителями» случившегося — «подглядывавшими» третьими особями, и закономерно модифицировать сигнальное поле группировки, всё равно запаховое, визуальное или акустическое. Всё это формирует социальную среду, которая равным образом связывает «всех со всеми», кооперирует поведение индивидов, вполне независимых и «эгоистичных» до вступления в социум, принуждая к предсказуемой форме взаимодействий в предсказуемые моменты времени и в предсказуемых частях участка.
Эта предсказуемость и есть главный ресурс, движущий прогрессивное развитие социальности — как поддерживающих её сигнальных средств, в сторону ещё большей ритуализации, так и поддерживаемых ею отношений — в сторону ещё большего управления индивидами. Чем плох социобиологический взгляд на социальность с коммуникацией — тем что он не заметил всех этих вещей, а они здесь самые главные. См. «Как социобиология сама себя орицает. Ч.2«.
Или, например, как пишет А.В.Чабовский:
«В эволюционных схемах социальность рассматривают как набор частных адаптации к одному или нескольким экологическим факторам: прессу хищников или конкурентов, распределению ресурсов, климату, — обеспечивающих повышение приспособленности особей в конкретных экологических условиях (Alexander, 1991; Cahan et al., 2002). Поиск частных экологических коррелятов изменчивости социальных отношений относит на второй план собственно социальную среду как набор экологических и эволюционных факторов, требующих соответствующих адаптации (Гольцман и др., 1994). При этом, адаптации «одиночных» видов к условиям социальной среды остаются за рамками рассмотрения проблемы, хотя фундаментальное значение исследований их социального поведения для понимания процесса эволюции социальности общепризнанно (Waser, Jones, 1988; Radespiel, 1999).
Между тем, социальная среда — эта ближайший, наиболее стабильный и предсказуемый компонент окружающей среды особи [если она есть — у несоциальных видов её нет], и поэтому процесс отбора должен идти более эффективно, быстро и независимо от изменений менее предсказуемой и менее стабильной экологической среды (Гольцман и др., 1994).
Адаптация к условиям социальной среды может идти в двух направлениях (Гольцман и др., 1981; 1994; Попов, 1986; 1988; Попов, Чабовский, 1995; 2005): в условиях плотной стабильной и предсказуемой социальной среды — в направлении развития социальной зависимости, в частности приводить к выработке устойчивости к социальному стрессу, способности формировать и поддерживать социальные связи и включению плотностно-зависимых механизмов регуляции размножения; в условиях изменчивой и малопредсказуемой социальной среды — в направлении развития социальной индифферентности, исключающей формирование устойчивых связей и плотностно-зависимую регуляцию размножения.
В рамках этого подхода эволюционные изменения поведения, связанные с выбором той или иной стратегии адаптации к условиям социальной среды, проявляются скоррелированно на разных уровнях организации поведенческих процессов: от кинематики движений до динамики популяций (Goltsman, Borisova, 1993; Гольцман и др., 1994; Goltsman, Volodin, 1997; Попов, Чабовский 1995).
Этот подход фокусирует внимание на механизмах формирования и поддержания устойчивых связей, рассматривает социальную среду как самостоятельный эволюционный фактор и не предполагает «по умолчанию» направления эволюции социальности в сторону ее развития, но наоборот предлагает два направления адаптации в зависимости от насыщенности и предсказуемости социальной среды (Гольцман и др., 1994; Попов, Чабовский, 2005).
Плотность, предсказуемость социальной среды как экологического и эволюционного фактора зависит, как от самой социальной среды как среды самоформирующейся, так и от экологических или абиатических факторов, которые, определяя условия перцепции, влияют на характер социальных взаимодействий: их кинематику и динамику, вероятность и предсказуемость. В частности, в условиях открытых пространств, расширение потока стимулов должно способствовать выработке устойчивости к возрастающему социальному давлению и другим стрессирующим факторам и способности поддерживать социальные связи (Попов, 1986). Таким образом, в противовес традиционным подходам, не социальная среда выступает в роли адаптивного буфера между особью и внешней средой, а экологическая среда выступает в роли фильтра или модулятора социальной среды и, таким образом, определяет направление адаптации к ней формирующих ее особей…
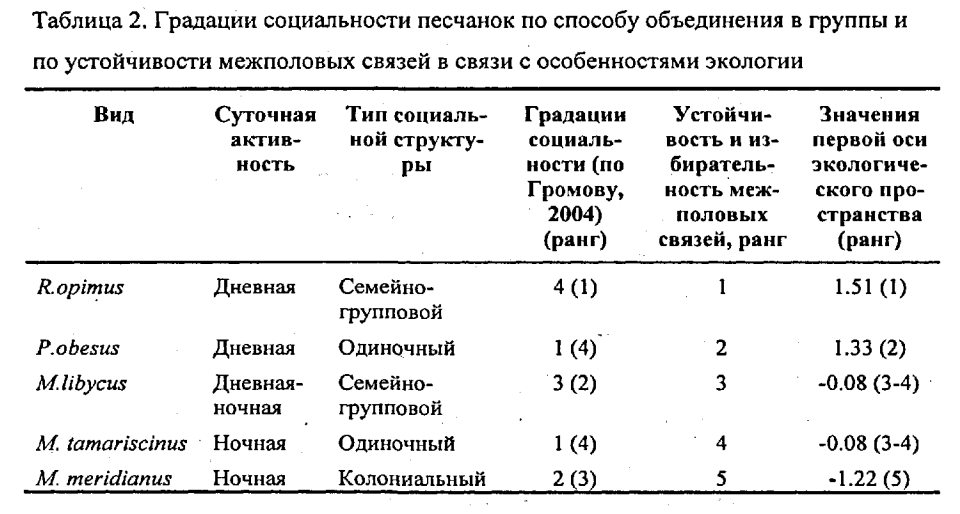
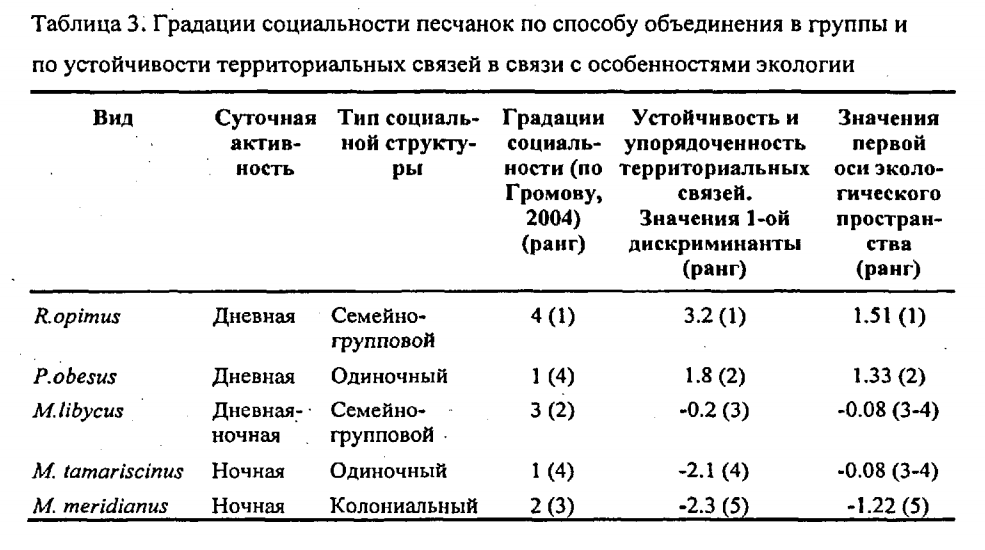
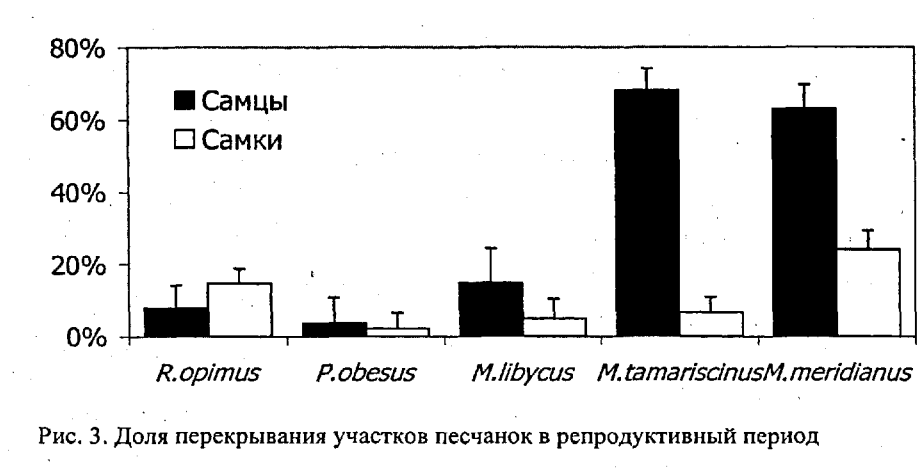
…В целом, количественный анализ упорядоченности и избирательности межполовых связей песчанок показывает следующее:
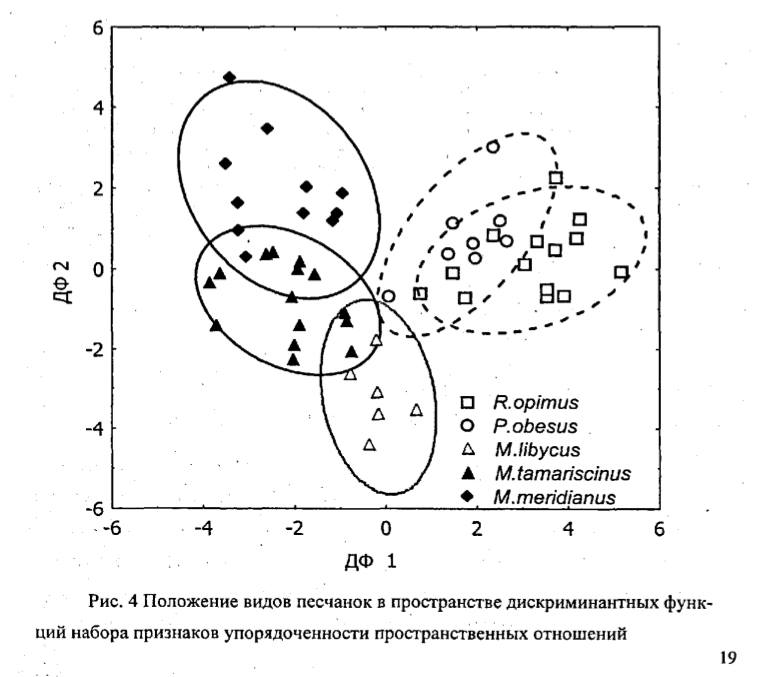
1. Шкала избирательности и упорядоченности межполовых связей как характеристики социальности не соответствует градациям социальности, выделяемым по типу социальной структуры (табл. 2): высокая степень упорядоченности связей может быть характерна, как для «высоко социальных» семейно-групповых видов», так и для «сугубо одиночных» песчанок. Низкая степень упорядоченности межполовых связей может быть характерна, как для «сугубо одиночных», так и для «более социальных» колониальных видов.
2. «Примитивные» в рамках традиционной схемы эволюции социальности грызунов одиночные P.obesus демонстрируют высокую степень упорядоченности межполовых связей и в этом аспекте наиболее близки к наиболее продвинутым в отношении «сложности» социальной структуры R.opimus. Колониальные M.meridianus, относимые к промежуточной стадии в эволюции социальности грызунов, демонстрируют наименьшую степень упорядоченности межполовых связей, меньшую, чем у «строго одиночных» M.tamariscinus.
3. Устойчивость межполовых связей в отличие от градаций социальности по составу групп хорошо коррелирует с экологической изменчивостью видов (табл.2, Kendall Tau=0.80, p=0.042) и увеличивается с переходом к дневной активности, зеленоядности, открытым местообитаниям и сложным норам, а также с размером тела (Kendall Tau=0.89, р=0.042). Наибольшая степень упорядоченности половых связей свойственна эволюционно продвинутым видам (R.opimus и P.obesus). ».
 Чабовский А.В., op.cit.
Чабовский А.В., op.cit.
Ну и последнее:
Размерный полиморфизм, как веса, так и «мозгов», авторами полностью игнорируется, что у видов — транспалеарктов делать никак нельзя. У них всех северные подвиды крупные, южные и/или горные существенно мельче, у больших пёстрых дятлов вес варьирует от 60 до 100 г., что между Северной и Южной Европой/Канарами, что между Евразией и Талышем/Китаем), авторы берут 80 г. — какого подвида? На каком основании? То же верно и для других видов с крупными ареалами и сильной подвидовой дифференциацией, скажем транссевероамериканскими, как у пушистопёрого дятла.

Свежие комментарии