 Print
Print  PDF
PDF
Бразильская свинка Cavia aperea, дикий предок домашней морской
В продолжение темы парфорсной эволюции
В биомедицинской науке последствия стресса в раннем онтогенезе в большинстве случаев рассматриваются как негативные для организма, вызывающие нарушения развития репродуктивных функций, иммунной, нейроэндокринной систем, мозга и поведения, включая когнитивные способности и устойчивые расстройства психики. На этом фоне набирают силу исследования, обсуждающие последствия ранних стрессов с позиции их возможной адаптивности, оцениваемой как увеличение приспособленности (fitness) в текущем и последующих поколениях. В обзоре дано краткое описание основных механизмов обусловленного материнским стрессом онтогенетического программирования организма потомка на основе обобщающих работ по млекопитающим. Основное же внимание уделено экспериментам в природе, либо экспериментам, моделирующим природную ситуацию, в которых на некоторых видах позвоночных животных были исследованы последствия материнских стрессов для потомков на фоне контролируемых изменений среды обитания, включая и среду социальную. Фенотипически проявляющиеся у потомков последствия ранних стрессов оказываются полезными для них (адаптивными) в тех случаях, когда условия среды (в т. ч. социальной), в которой существовал организм матери в период формирования яйца или беременности и сразу после рождения выводка, соответствуют условиям среды их будущей жизни. Для матери стрессовые состояния, транслируемые потомкам, могут быть выгодны, поскольку оптимизируют затраты на текущее размножение и в конечном счете могут повышать ее совокупную приспособленность независимо от того, повышают они или понижают приспособленность потомков в ближайшей перспективе. Эволюционно-экологический подход в изучении последствий ранних стрессов может быть полезным для понимания процессов формирования здоровья человека. Накопленные на сегодня данные вселяют надежду на то, что будет наконец достигнуто понимание необходимости сокращения несоответствия между пренатальной и будущей средой жизни человека, несоответствия, существенно повышающего риски заболеваний в зрелом возрасте.
В океане исследований стресса периода раннего онтогенеза и его последствий существует, однако, относительно молодое и обособленное течение – исследования, обсуждающие последствия ранних стрессов с позиции их эволюционной значимости. Стрессовые состояния материнского организма до рождения детеныша или откладки яйца и стрессоры, воздействующие на новорожденный организм в период, непосредственно следующий за появлением на свет, влияют на его развитие, отражаются на строении формирующихся органов и систем, механизмах функциональных связей и поведении (Maccari et al., 2003; Weinstock, 2005; Lupien et al., 2009; McEwen, 2012; Sheriff, Love, 2013).
В ходе формирования в онтогенезе физиологических функций и механизмов поведения происходит их настройка – “программирование” – в соответствии с текущими и ожидаемыми условиями жизни организма. Фенотипические эффекты этих изменений могут проявляться в течение жизни не только как различные отклонения от нормы, снижающие приспособленность, но и как вполне адаптивные явления, значение которых становится понятным при рассмотрении их на фоне меняющихся условий жизни в контексте конфликтов жизненного цикла (life history tradeoffs) и событий популяционного уровня (Sheriff et al., 2011; Sheriff, Love, 2013). Сам же механизм онтогенетического программирования может иметь адаптивную эволюционно-генетическую основу и обеспечивать передачу эффекта неблагоприятного воздействия, перенесенного предками, в ряду поколений (Дыгало, 2007).
Научная литература, посвященная последствиям для потомства стресса, который испытывала мать (matternaly derived stress), как и вообще стрессу в раннем онтогенезе, поистине огромна. Публикации на эту тему исчисляются тысячами. В биомедицинской науке последствия стресса в раннем онтогенезе в большинстве случаев рассматриваются как негативные для организма, вызывающие нарушения развития репродуктивных функций (Dobson, Smith, 2000), иммунной (Merlot et al., 2008) и нейроэндокринной систем (Glover et al., 2010), мозга и поведения, включая когнитивные способности и устойчивые расстройства психики (шизофрения, депрессия, синдром повышенной тревожности, аутизм и др.; Weinstock, 2008; Charil et al., 2010; Glover, 2011; Shishkina, Dygalo, 2017). Этим обусловлен непрекращающийся интерес к проблематике.
Влияние повышенной адренокортикальной активности (стрессированности) на приспособленность (fitness) к настоящему времени хорошо документировано (Dufty et al., 2002; Cavigelli, McClintock, 2003; Boonstra, 2005; Wingfield, 2005; Hadany et al., 2006; Blas et al., 2007; Роговин, Мошкин, 2007; Новиков, Мошкин, 2009). Адаптивное значение внутри- и межпопуляционной изменчивости в уровне стрессированности неоднократно подтверждалось исследованиями в природе.
Это и снижение чувствительности к стрессорам у размножающихся в горах и в Арктике птиц (Wingfield, Hunt, 2002; Breuner, Hahn, 2003), и снижение реактивности гипоталамо-гипофизарно-надпочечниковой системы к стрессорам в течение репродуктивного цикла, уменьшающее вероятность его прерывания (Holberton, Wingfield, 2003; Love et al., 2004), и коррелирующие с уровнем глюкокортикоидной активности выживаемость и вероятность рекрутирования молодых животных в локальную популяцию, социум (Cavigelli, McClintock, 2003; Blas et al., 2007; Meylan et al., 2012).
В основе этих адаптаций мог лежать механизм онтогенегического программирования. Число исследований эволюционной направленности последствий стресса в раннем онтогенезе невелико и по сравнению с классическим периодом эволюционной экологии, с присущим этому периоду пристальным вниманием к стрессу как регулятору популяционных процессов (Christian, 1950, 1963, 1968, 1971, 1980; Шилов, 1984; Мошкин и др., 1990; Wingfield, Ramenofsky, 1999; Wingfield, Romero, 2001; Роговин, Мошкин, 2007; Новиков, Мошкин, 2009). Между тем именно популяционный аспект проблемы стресса в раннем онтогенезе представляется чрезвычайно интересным не только в плане теоретическом, но и в плане возможности прогнозирования демографических последствий стрессов, испытываемых в раннем онтогенезе человеком (Glover, 2011).
Вопрос о существовании адаптивной компоненты явления не имеет смысла вне связи с условиями его реализации. Однако среди многочисленных исследований влияния материнского стресса на развитие структур и функций мозга, кортикостероидной оси стресса и поведения потомков, лишь в немногих из них эффекты материнского стресса оцениваются с учетом условий пренатального периода и постнатальной жизни (Blas et al., 2007; Cyr, Romero, 2007; Lowe, Williams, 2008a, b; Sheriff et al., 2011; Sheriff, Love 2013).
Предлагаемый вниманию читателей обзор посвящен обсуждению адаптивного значения последствий стресса в раннем онтогенезе. В обзоре будет дано краткое и, в силу этого, неизбежно поверхностное описание механизма, обусловленного материнским стрессом и основанного на геномных взаимодействиях онтогенетического программирования организма потомка на основе обобщающих работ по млекопитающим. Эта тема огромна и выходит за рамки нашей задачи. Основное внимание будет уделено экспериментам в природе, либо экспериментам, моделирующим природную ситуацию, в которых на некоторых видах позвоночных животных были исследованы последствия материнских стрессов для потомков на фоне контролируемых изменений среды обитания, включая и среду социальную (социальное окружение).

Ганс Селье
Теория стресса была сформулирована Селье (Selye, 1950, 1952, 1981) применительно к организму млекопитающего. Механизм неспецифического адаптационного синдрома (Selye, 1950, 1952, 1981) был подробно изучен и исследуется ныне главным образом у немногих модельных видов млекопитающих (крысы, мыши, морские свинки, некоторые обезьяны), а также у человека (Wingfield, Sapolsky, 2003; Weinstock, 2008; McEwen, 2012; McEwen et al., 2012). Однако механизм нейроэндокринных взаимодействий кортикоидной оси стресса консервативен. В основных чертах он присущ всем позвоночным животным (птицы: Banerjee et al., 2012; Zimmer, Spencer, 2014; рептилии: Moore, Jessop, 2003; Silvestre, 2014; амфибии: Moore, Jessop, 2003; Hu et al., 2008; современные костистые рыбы: Feist, Schreck, 2001; Auperin, Geslin, 2008; Steenbergen et al., 2011).
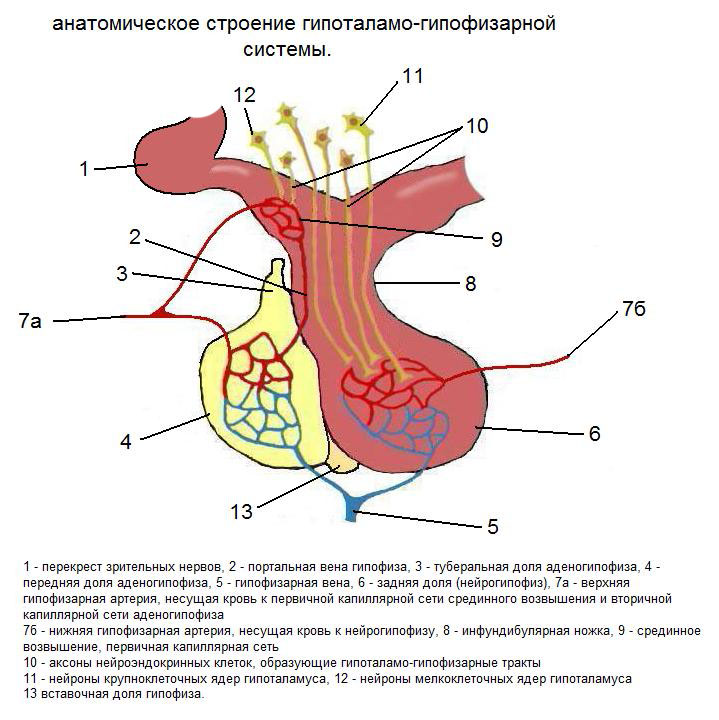
У амниот действует гипоталамо-гипофизарно-надпочечниковая система, конечным продуктом которой и основным регулятором являются выделяемые корой надпочечников глюкокортикоиды – кортизол или кортикостерон (Sapolsky et al., 2000; Wingfield, Sapolsky, 2003; Wingfield, 2005). У костистых рыб и амфибий действует гипоталамо-гипофизарно-интерренальная система, в которой глюкокортикоидный гормон вырабатывается интерренальными железами (Wendelaar Bonga, 1997; Bernier, Peter, 2001; Dores, Garcia, 2015).
Принятые в статье сокращения
ГГНС (HPA – hypothalamic-pituitary-adrenal axis) – гипоталамо-гипофизарно надпочечнико-вая система (ось). Мы используем традиционную аббревиатуру ГГНС, хотя в действительности помимо собственно гипоталамуса, гипофиза и надпочечников в центральную нейроэндокринную ось стресса у млекопитающих входит почти вся лимбическая система мозга и фронтальная кора, где имеются рецепторы кортикоидных гормонов (McEwen, 2012); ГК – глюкокортикоиды; МК – минералокортикоиды; ГР – рецепторы глюкокортикоидов, продукты активности генов (gr); МР – рецепторы минералокортикоидов, продукты активности генов (mr); КРГ (CRH – corticotropin-releasing hormone) – кортикотропин-рилизинг гормон, кортиколиберин; СТМ (МDS – maternally derived stress) – стресс, транслированный организмом матери организму потомка.
Границы понятия “стресс в раннем онтогенезе”
Содержание
Под стрессом в раннем онтогенезе мы понимаем нейроэндокринные эффекты, которые организм матери с повышенной активностью центральной нейроэндокринной оси стресса оказывает на созревающий ооцит или плод, и меняющие его развитие с длительными относительно устойчивыми последствиями, проявляющимися в течение жизни потомка. Сюда же относятся эффекты, обусловленные родительским поведением матери, социальным окружением, хищником или действием других внешних стрессоров в ранний постнатальный период жизни, влияющие на ход нормального развития организма потомка. Стресс в раннем онтогенезе близок к часто используемому понятию стресса, транслируемого организмом матери потомкам (maternally derived stress).
В исследованиях СТМ у млекопитающих внимание акцентируется на повышенной активности ГГНС матери и гуморальных эндокринных влияниях материнского организма на развивающийся плод (пренатальный стресс у млекопитающих). В понятие СТМ также входят поведенческие эффекты матери (качество материнской заботы) в неонатальный период (ранний постнатальный период). У яйцекладущих животных влияние стресса матери на потомство распространяется на весь период от развития фолликула до откладки яйца и период родительской заботы (у птиц, некоторых рыб). В понятии СТМ, однако, не учтена возможность прямых стрессирующих воздействий на уже рожденных детенышей социального окружения, хищников и пр. (Если они оцениваются, то опосредованно через изменение поведения матери).
О перинатальном стрессе говорят применительно к млекопитающим; рассмотрение ограничивается периодом, непосредственно предшествующим родам (поздним пренатальным), и коротким, следующим сразу за ними (неонатальным). У человека перинатальный (околородовой) период исчисляется, начиная с 22-й полной недели (154-го дня) внутриутробной жизни плода по 7-й день после рождения ребенка.
Этими ограничениями и объясняется использование нами сравнительно редко употребляемого словосочетания “стресс в раннем онтогенезе”. Именно этот термин наиболее полно очерчивает круг явлений, представляющихся значимыми в попытке интерпретировать эффекты ранних стрессов с позиции их адаптивности.
Стрессоры в моделях изучения эффектов стресса в раннем онтогенезе
Повышенная анормальная активность ГГНС матери в период до рождения потомства или откладки яйца может быть обусловлена как различными внешними воздействия на материнский организм, так и внутренними причинами – устойчивыми болезненными состояниями матери. К стрессорам раннего постнатального периода относятся как внешние по отношению к системе “мать–потомство” негативные факторы, с которыми сталкиваются появившиеся на свет детеныши, так и измененное поведение матери (родителей), в первую очередь нарушенная (недостаточная) забота о потомстве. В экспериментальных исследованиях пренатального стресса обычно используют стандартизированные стрессоры, такие как иммобилизация (Fujioka et al., 1999; Darnaudéry, Maccari, 2008), иммобилизация в комбинации с ярким освещением (Frye, Orecki, 2002), звук (Coe et al., 1999), звук в комбинации со вспышками света (Fride, Weinstock, 1988), удар током (Fonseca et al., 2002), взятие в руки (Braastad et al., 1998) или более экзотические воздействия, такие как лишение сна (Suchecki, Palermo-Neto, 1991) или погружение в холодную воду (Drago et al., 1999). Стандартизация воздействий важна при исследовании тонких механизмов программирования мозга, ГГНС, поведения. Однако эти стрессоры не имеют ничего общего с реальными ситуациями, с которыми животные сталкиваются в природе. Более реалистичны стрессоры социальной природы: помещение беременной самки в группу незнакомых животных (Coe, Crispen, 2000), периодические ссаживания с незнакомыми беременными самками (Marchlewska-Koj et al., 2003a, b), поражение при ссаживании на нейтральной территории (Stefanski et al., 2005), помещение в социальную среду с высокой плотностью (Crump, Chevins, 1989), добавление самцов и самок в группу (Zielinski et al., 1991), уплотнение с перемещением животных из группы в группу (Harvey, Chevins, 1987).
Все эти манипуляции помимо эффектов прямых (не только агрессивных) взаимодействий означают нарушение стабильности социальной среды, резко увеличивают неопределенность, непредсказуемость ситуации, что само по себе является мощным фактором стресса (Von Holst, 1998). Нарушение стабильности социальной среды перемещением животных из группы в группу было успешно использовано при изучении последствий пренатального стресса в серии экспериментальных работ на морских свинках (Kaiser, Sachser, 1998, 2001; Kaiser et al., 2003а, b).
В исследованиях раннего постнатального стресса в качестве стрессоров используют разные варианты ранней социальной депривации: у приматов отлучение от матери и (или) от других членов группы (Harlow et al., 1971; Suomi, 1991; Winslow et al., 2003), у грызунов – различные формы изоляции от матери в сочетании со взятием в руки (различные варианты постнатальной социальной депривации у крыс и мышей подробно рассмотрены в статье Виинема (Veenema, 2009)).
В природе наиболее вероятными и наиболее изученными факторами стресса в период раннего онтогенеза являются хищники и обеспеченность ресурсами. Риск быть пойманным хищником увеличивается с ростом численности хищника и ведет к увеличению уровня адренокортикальной активности в организме самок (млекопитающие: Boonstra et al., 1998; Monclús et al., 2011; Sheriff et al., 2009, 2011; птицы: Saino et al., 2005; Love et al., 2008; Zanette et al., 2011; рыбы: McCormick, 1998; Giesing et al., 2011). Увеличение уровня стрессированности самок может быть прямым образом связано с уменьшением предсказуемости, качества и количества пищевых ресурсов (Kitaysky et al., 2007; Rogovin et al., 2008; Shultz, Kitaysky, 2008; Welckler et al., 2009).
Повышенный уровень ГК самки может быть опосредован ухудшением ее состояния в связи с недостаточностью питания (Fraipont et al., 2000; Meylan et al., 2002; Love et al., 2009). Возможен и синергетический эффект взаимодействия факторов обеспеченности пищей и хищничества (Sheriff et al., 2010). Социальные факторы также могут выступать в качестве стрессоров в природе. Это стресс, обусловленный ростом локальной плотности (классическая модель Кристиана – Christian, 1963, 1968, 1971, 1980) и внутригрупповыми отношениями (Sapolsky et al., 2000; Creel, 2001; Creel et al., 2013; Young et al., 2006). Агрессивные взаимодействия в конкурентной среде ведут к росту уровня ГК в организме самок (рыбы: МсCormick, 2006; пресмыкающиеся: Comendant et al., 2003; птицы: Love et al., 2008; млекопитающие: Kaiser, Sachser, 2005). В природной обстановке стрессорами, негативно влияющими на материнский организм, могут быть и климатические факторы (Wingfield, 2008), и влияние человека (Thiel et al., 2008; Wasser et al., 2011).
В природе факторы, обусловливающие состояние материнского организма, действуют совместно. В ранний постнатальный период нарушения поведения самки (родителей), вызванные ее состоянием, недостатком пищевых ресурсов, присутствием хищников и пр., являются фактором стрессированности потомков, наряду с прямыми негативными воздействиями на них среды.
В этом случае, однако, нельзя разделить следствия, связанные с недостаточностью питания или заботы, и прямые эффекты биотических и физических стрессоров (Sheriff, Love, 2013).
Стресс и эпигенетическое программирование организма в раннем онтогенезе
Гормоны ГГНС играют важную роль в эпигенетическом программировании развития организма млекопитающего. В первую очередь такое программирование касается структурной организации головного мозга (McEwen, 2012). Мозг млекопитающих активно реагирует на хронические стрессовые нагрузки; в той или иной форме изменения отмечаются почти во всех отделах лимбической системы, включая поясную и зубчатые извилины, гиппокамп, миндалевидное тело (amigdala), гипоталамус (Hunter, 2012). На модельных видах грызунов (мыши, крысы) показано, что в области гиппокампа и медиальной префронтальной коры хронический стресс вызывает уменьшение длины дендритных окончаний, их ветвления и как следствие – сокращение площади синапсов. В этом, наряду с эффекторными аминокислотными трансмиттерами (гамма аминомасляная кислота, глицин и глутаминовая кислота), принимают самое непосредственное участие гормоны стресса – глюкокортикоиды. В отличие от гиппокампа и префронтальной коры хронический стресс ведет к росту дендритных окончаний и увеличению синаптических связей в базолатеральной области миндалины и орбитофронтальной области коры, что обусловливает рост тревожности, настороженности и агрессивности (McEwen, 1999, 2007).
 Такие изменения обратимы, однако вероятность этого по выходе из стрессового состояния уменьшается с возрастом (Bloss et al., 2010). Сходные изменения возможны и вследствие стресса, который организм может испытывать в раннем онтогенезе (Weinstock, 2001, 2005, 2008). В развивающемся мозге структурные изменения в ответ на пренатальный стресс наблюдаются в префронтальной коре, миндалевидном теле, гиппокампе и гипоталамусе. Мозг запечатлевает стрессовые влияния эпигенетически путем включения механизмов метилирования цитозиновых оснований в промоторной зоне генов – рецепторов гормонов коры надпочечников (адренокортикоидов), либо путем модификации гистонов – белков, обслуживающих процесс транскрипции, или путем регуляции процесса трансляции мРНК малыми некодирующими молекулами РНК (Hunter, 2012; Hunter, McEwen, 2013).
Такие изменения обратимы, однако вероятность этого по выходе из стрессового состояния уменьшается с возрастом (Bloss et al., 2010). Сходные изменения возможны и вследствие стресса, который организм может испытывать в раннем онтогенезе (Weinstock, 2001, 2005, 2008). В развивающемся мозге структурные изменения в ответ на пренатальный стресс наблюдаются в префронтальной коре, миндалевидном теле, гиппокампе и гипоталамусе. Мозг запечатлевает стрессовые влияния эпигенетически путем включения механизмов метилирования цитозиновых оснований в промоторной зоне генов – рецепторов гормонов коры надпочечников (адренокортикоидов), либо путем модификации гистонов – белков, обслуживающих процесс транскрипции, или путем регуляции процесса трансляции мРНК малыми некодирующими молекулами РНК (Hunter, 2012; Hunter, McEwen, 2013).
Хотя эндокринное влияние матери у млекопитающих не исчерпывается гормонами ГГНС, их влияние изучено в наибольшей степени (McEwen, 2012; McEwen et al., 2012; Hunter, McEwen, 2013). Ниже мы лишь кратко охарактеризуем программирование мозга, ГГНС и поведения в пренатальный период, обусловленное влиянием гормонов ГГНС матери, а также программирование фенотипических качеств потомков в ранний постнатальный период, в котором ведущую роль играют родительские качества матери. Обсуждению механизмов программирования фенотипических особенностей потомства (морфологических, физиологических, поведенческих) посвящен ряд специальных обзоров (Seckl, 2004; Seckl, Meaney 2004; Meaney et al., 2007; Darnaudéry, Maccari, 2008; Weinstock 2001, 2005, 2008; McEwen, 2012; McEwen et al., 2012; Hunter, McEwen, 2013, и др.).
Пренатальное программирование
Существует большое число исследований лабораторных млекопитающих, свидетельствующих о влиянии материнского стресса (в основном на поздних стадиях беременности) на функционирование ГГНС и поведение потомков в зрелом возрасте (Schneider et al., 2002; Matthews et al., 2004; Kloet et al., 2005; Owen et al., 2005; Meaney et al., 2007; Weinstock, 2001, 2008). Гормоны стресса – глюкокортикоиды (ГК) играют важную роль в развитии мозга. Они взаимодействуют с внутриклеточными рецепторами двух типов, с высоко аффинными рецепторами глюкокортикоидов (ГР) и низкоафинными к ГК рецепторами минералокортикоидов (МР). Эти рецепторы действуют как лигандактивируемые транскрипционные факторы (Kloet et al., 1998). Активация ГР происходит с повышением уровня ГК в состоянии стресса и зависит от стадии суточного цикла (утром у людей, вечером и ночью у мышей и крыс). ГР широко представлены в разных отделах мозга – во фронтальной и поясной коре, гиппокампе, базолатеральном и базомедиальных ядрах миндалины, в таламусе, но особенно многочисленны в ядрах гипоталамуса и в гипофизе, где на них замыкаются основные контуры обратных связей ГГНС. Наиболее высокая концентрация МР характерна для гиппокампа (Kloet et al., 1998; Matthews, 1998). Избыточная экспозиция ГК или, наоборот, их недостаточность в период эмбриогенеза (модуляция в эксперименте) ведут к структурным и функциональным изменениям в мозге и в поведении (Sapolsky, 1987; Muneoka et al., 1997; Matthews, 2002). Так, показано, что пренатальная экспозиция ГК ведет к уменьшению представительства ГР и МР в гиппокампе, ослабляя механизм действия обратной связи ГГНС (Maccari et al., 1995, Van Waes et al., 2006). С этим связывают повышенный уровень ГК в организме взрослых потомков (Henry et al., 1994; Maccari et al., 1995; Levitt et al., 1996; Welberg et al., 2001; Welberg, Seckl 2001; Emack et al., 2008).
У млекопитающих в норме развивающийся плод защищен от эндогенных материнских ГК плацентарным барьером. У людей кортикотропин-релизинг гормон (КРГ) вырабатывается не только в мозге, но и плацентой (отличие приматов от грызунов, у которых плацентарный КРГ отсутствует; Weinstock, 2008). У беременных женщин плод оказывается защищенным от избытка материнского КРГ (Perkins et al., 1995). Уровень КРГ в плазме крови матери увеличивается в несколько раз в течение последнего триместра беременности, но гормон остается неактивным до тех пор, пока он соединен со связывающим белком (CRH-BP), уровень которого падает лишь к моменту родов. В результате увеличивается концентрация свободно циркулирующего КРГ, под действием которого ГГНС матери и плода продуцируют АКТГ и β-эндорфин (Chan et al., 1993).
В норме концентрации КРГ, АКТГ и β-эндорфина в плазме крови повышены в начале третьего триместра беременности (Wadhwa et al., 1996; Weinstock, 2005). Концентрация кортизола в крови матери также постепенно увеличивается и к концу третьего триместра беременности в 2–3 раза превышает таковую у небеременных. При этом концентрация кортизола в крови плода оказывается в 13 раз ниже, чем у матери (Gitau et al., 2001). Происходит это потому, что 80% его метаболизируется 11β-гидроксистероиддегидрогеназой-2 (11β-HSD2) до состояния неактивного кортизона в плаценте. 11β-HSD2 способна конвертировать и кортикостерон в инертный 11-дегидрокортикостерон, DH-B (Funder, 1996; Burton, Waddell, 1999; Seckl, 2004). Однако, в том случае, когда мать испытывает сильное воздействие стрессора, экспрессии 11β-HSD2 не происходит, либо она ослабевает (Lucassen et al., 2009). В результате развивающийся организм оказывается неспособным противостоять стрессирующему влиянию материнского организма.
Конкретные механизмы влияния материнских ГК на развитие мозга потомка все еще недостаточно исследованы. Хотя прямые мутагенные эффекты также существуют, эти механизмы в основном эпигенетические, определяющие изменения в экспрессии генов без изменения самих нуклеотидных последовательностей (Szif et al., 2008; McGowan, Szif, 2010a,b). Наиболее частым механизмом является метилирование ДНК и, как следствие, транскрипционные нарушения (Mueller, Bale, 2008; Peña et al., 2012; Matrisciano et al., 2013). Надежно показано, что обусловленное пренатальным стрессом програмирование мозга ориентировано по полу (Weinstock, 2007). Так, гиперметилирование цитозиновых оснований в области промотора ГР 1-7 в гиппокампе и гипометилирование рецептора КРГ в гипоталамусе и центральной зоне миндалины было обнаружено у подвергавшихся пренатальному стрессу самцов мышей, но не наблюдалось у самок. Эти различия в реакции полов коррелировали с экспрессией ряда генов в плаценте (Mueller, Bale, 2008).
Другой механизм связан с участием miRNA – малых некодирующих молекул РНК, влияющих на процесс трансляции. Показано, что эти эффекты сохраняются и во втором поколении потомков (Morgan, Bale, 2011).
Дифференцированные по полу эффекты материнского стресса подробно исследованы на крысах. Пренатальный стресс вызывает повышение уровня кортикостерона в эмбриональном мозге самцов, ведет к феминизации мозга и поведения. Под действием кортикостерона в развивающемся мозге самцов крыс снижается концентрация тестостерона и фермента ароматазы (обеспечивает превращение тестостерона в эстрадиол), меняется активность катехоламинов, происходит уменьшение размера диморфного по полу ядра преоптической области (SDN-POA) и истончение отдельных участков коры.
Проблемы с обучаемостью, замедление нейрогенеза и долгосрочной симпатической трансмиссии (hippocampal long termed potentiation, LPT) в гиппокампе, редукция плотности дендритных шипиков в префронтальной коре более выражены у пренатально стрессированных самцов, в то время как синдромы тревожности, депрессии и повышенная реактивность ГГНС к воздействию стрессоров более выражены у пренатально стрессированных самок. Гендерные различия существуют в чувствительности развивающегося мозга к гормонам стресса (Weinstock, 2007). В целом у грызунов гендерные различия в структуре и функционировании мозга и поведении в результате влияния материнских ГК сохраняются и во взрослом состоянии, но в измененном виде (Catalani et al., 2011).
Вейнсток (Weinstock, 2005, 2008) подробно проанализировала факторы, с которыми может быть связана вариабельность результата пренатального программирования качеств потомства у грызунов. У крыс и мышей каждый из факторов, таких как природа и время экспозиции стрессора во время беременности, возраст, в котором проводится тестирование потомков в отношении эффектов материнского стресса, генетические особенности исследуемых линий, а также пол потомка, обусловливал фенотипические эффекты испытанного в пренатальный период стресса. К этому необходимо прибавить влияние на результат времени суток, в которое проводилось тестирование ГГНС потомков.
Результат оценки материнского эффекта зависел от того, исследовали ли у потомков фоновый уровень глюкокортикоидов или уровень глюкокортикоидов в ответ на воздействие стрессора. Кроме того, существует вариабельность пороговых уровней реагирования развивающегося плода на стресс матери как на индивидуальном, так и на видовом (генетические линии) уровнях. Для возникновения эффекта воздействие стрессора должно быть достаточно длительным и интенсивным. Как правило, последствия наиболее выражены, если материнский организм подвергался воздействию стрессора на поздней стадии беременности. Это связано со временем формирования структур мозга, рецепторов и системы функциональных связей в мозге и организме в целом. У разных видов эти структуры и связи формируются в разное время. С этим связана видоспецифичность эффектов материнского стресса (Weinstock, 2008).
Таким образом, обусловленное материнским стрессом пренатальное программирование потомства у млекопитающих (по-видимому, подобная схема существует и у других позвоночных) включает цепь событий от влияния стрессоров на организм матери, эндокринных взаимодействий организма матери с организмом плода с участием плаценты, до эпигенетических изменений в организме потомка. Материнское неблагополучие в период беременности ведет к повышению материнского ГК сигнала, к уменьшению экспрессии ГР у развивающегося плода и к повышенному уровню ГК у потомка. Соответственно меняется порог реагирования организма потомка на стрессоры.
Постнатальное программирование
Степень завершенности развития (созревания) мозга и ГГНС относительно даты рождения различается у разных модельных видов млекопитающих. У приматов, жвачных, морских свинок формирование мозга и нейроэндокринной системы заканчивается в основном к моменту рождения. Однако у мышей, крыс, кроликов эти процессы продолжаются и в раннем постнатальном (неонатальном) периоде жизни (Weinstock, 2008). Еще в 50-е годы ХХ в. стало известно, что многократное взятие в руки детенышей крыс и мышей в отдельные периоды неонатального развития (действие, моделирующее усиленную родительскую заботу) снижает стрессируемость в ответ на воздействие стрессоров в последующей жизни (Levine, 1957, 1960; Denenberg, Karas, 1959).
С тех пор накоплено большое количество свидетельств тому, что степень связи с матерью, ее заботливость в ранний постнатальный период влияет на экспрессию рецепторов глюкокортикоидов, чувствительность к стрессорам и изменяет поведение потомков в стрессовых ситуациях (Francis, Meaney, 1999; Meaney, 2001; Meaney et al., 2007). Так, подросшие потомки самок крыс, больше заботившихся о детенышах, демонстрировали во взрослом состоянии повышенную экспрессию ГР в гиппокампе, отличались повышенной функциональной активностью контура отрицательной обратной связи ГГНС и имели более сдержанный по сравнению с контролем ответ на воздействие стрессоров (Liu et al., 1997). Во взрослом состоянии такие потомки демонстрировали высокий уровень заботы о детенышах. Экспериментальные манипуляции с перекладыванием детенышей от заботливых матерей к незаботливым и наоборот показали, что замена родной матери на чужую в первый день постнатальной жизни реверсивно меняет фенотип детенышей в соответствии с тем, какая самка воспитывает детенышей.
Это свидетельствует о прямой связи тренда развития ГГНС и поведения потомков с качеством материнской заботы (Francis et al., 1999; Meaney, 2001). Эксперименты с перекладыванием (cross-fostering) детенышей также показали, что именно стресс в пренатальный период обусловливает изменения у потомков.
В условиях постнатального стресса наиболее частым механизмом регуляции активности генов является метилирование ДНК, однако наряду с этим описан механизм, основанный на модификации гистонов – белков, обслуживающих процесс танскрипции (Hunter, McEwen, 2013). Так, материнская забота способна влиять на степень метилированности ДНК в промоторе гена ГР в гиппокампе (Weaver et al., 2004). Это происходит в результате ингибирования байндинг-активности транскрипционного фактора NGFI-A, обеспечивающего экспрессию ГР (Weaver et al., 2004, 2005, 2007). Присутствие метилированных нуклеотидов в местах связывания узнаваемых NGFI-A транскрипционных факторов с нуклеотидной последовательностью ГР-промотора вызывает остановку транскрипции под действием РНК-полимеразы (наблюдается снижение экспрессии мРНК). Нуклеотидные последовательности в зоне промотора ГР характеризуются более низким уровнем метилированности у потомков, матери которых были более заботливыми, и наоборот уровень метилированности ДНК был более высоким у потомков плохих матерей. Более высокий уровень метилированности ДНК в зоне промотора гена ГР обусловливал снижение количества ГР, приводил к меньшей стресс-реактивности и более медленному восстановлению после прекращения действия стрессора у взрослых животных.
Эти различия, проявившиеся уже на первой неделе жизни, оказались, однако, обратимыми, когда детенышей от заботливой матери перекладывали к малозаботливой (cross-fostering experiment), и наоборот. В дальнейшем изменения сохранялись во взрослом состоянии. Если же в мозг детенышей незаботливых матерей вводили трихостатин А (ингибитор гистондиацетилазы, обеспечивает более открытую конфигурацию хроматина и способен интенсифицировать транскрипцию), а в мозг детенышей заботливой самки вводили метионин (донор метильной группы, усиливающий метилирование ДНК в присутствии метилтрансферазы), различия между группами по уровню метилированности ДНК, связанности NGFI-A с промотором ГР, ГР-экспрессии и реактивности ГГНС в ответ на стрессоры исчезали.
То, что “хорошее” материнское поведение может компенсировать негативные последствия пренатального стресса, известно из других исследований. Беременные самки крыс, подвергавшиеся воздействиям стрессоров на последней неделе беременности, демонстрируют ухудшенное материнское поведение выкармливания. Однако если сравнить стрессированных потомков крыс, выращенных нестрессированными матерями, с нестрессированными потомками, выращенными стрессированными во время беременности самками, то оказывается, что материнская забота со стороны нестрессированных матерей способна компенсировать негативное влияние пренатального стресса на родительские качества дочерей вне связи с гормональными или нейрональными перестройками в период эмбрионального развития.
Таким образом, не только пренатальный стресс, но и качество материнской заботы в ранний постнатальный период влияют на поведенческий фенотип потомка (Del Cerro et al., 2010). Свидетельства эпигенетической регуляции в мозге в связи с качеством родительской заботы и другими стрессирующими воздействиями в ранний постнатальный период жизни получены и другими группами исследователей (Murgatroyd et al., 2009; Roth et al., 2009; McGowan et al., 2011). Одним из стрессирующих факторов может выступать социальное окружение, влияющее на нейрогенез в мозге мышей (Branchi et al., 2006). Экспериментальные исследования свидетельствуют о том, что особенности взаимодействия детенышей с матерью в ранний постнатальный (неонатальный) период жизни играют важную роль в программировании функциональных связей мозга, ГГНС, способности реагировать на стрессирующие воздействия среды в течение взрослой жизни. Высокое качество материнской заботы, связанное часто с низким уровнем стрессированности матери, обусловливает надежное функционирование контуров обратных связей ГГНС, замкнутых на богато представленные в структурах мозга рецепторы глюкокортикоидов, и, соответственно, обусловливает низкий уровень стрессированности у потомков.
Популяционные последствия стресса в раннем онтогенезе
1. Классический период популяционной экологии.
Гипотеза материнского эффекта и популяционные циклы численности Гипотеза авторегуляции, основанная на физиологических эффектах в сфере отношений матери и потомства, была высказана еще в 80-е годы прошлого века (Ward, 1984; Lee, McDonald, 1985).
В сущности она представляла собой расширительное толкование гипотезы обусловленной стрессом авторегуляции популяционной плотности Кристиана (Christian, 1950, 1963; Christian, LeMunyan, 1958) и предсказывала возможность регуляции популяционной плотности млекопитающих через определяемые условиями развития детенышей изменения их поведения в пре- и постпубертатном возрасте, в первую очередь в сфере репродуктивного поведения и пространственной активности (характер использования территории, способность к расселению). Предполагалось, что социогенный стресс (обусловленный избыточной плотностью социальной среды) может сказываться как на развитии плода, так и на развитии и темпах роста детенышей в постнатальный период, внося дополнительный вклад в снижение численности наряду с торможением репродуктивных функций и повышенной, обусловленной стрессом, смертностью.
Исследований, свидетельствующих о торможении роста, задержке созревания в условиях повышенной плотности населения существовало достаточно много (Marchlewska-Koj, 1997). Хорошим примером исследований этого периода может служить сравнительный анализ взаимозависимости между динамикой популяционной плотности, темпами полового созревания и адренокортикальной активности у красных полевок в оптимуме и пессимуме ареала (Евсиков, Мошкин, 1994; Novikov, Moshkin, 1998).
В оптимуме ареала на пике популяционной плотности было отмечено полное подавление полового созревания сеголеток и максимальная адренокортикальная активность у полевок разного пола и возраста. При содержании молодых животных, отловленных в начале лета, в отдельных клетках темпы созревания в сезоны максимальной популяционной плотности возрастали, а при низкой плотности в природе замедлялись (Novikov, Moshkin, 1998). Более детальные исследования с контролем формирования отдельных структур ГГНС были выполнены на копытных леммингах (Dicrostonyx torquatus), у которых на пике популяционной плотности было отмечено торможение пренатального и постнатального развития нейросекреторных ядер гипоталамуса, участвующих в регуляции гипофизарно-надпочечниковой и ги- пофизарно-гонадной систем (Аршавская, Данилова, 1987; Аршавская и др., 1987; Ткачев и др., 1987; Владимирова и др., 1996).
Высокие показатели эмбриональной и ювенильной смертности копытного лемминга, установленные на стадиях пика и спада численности (Ткачев и др., 1987), связывались с низким уровнем развития гипоталамо-гипофизарного комплекса. Предполагалось, что более раннее формирование механизмов гипоталамического контроля вегетативных и эндокринных функций должно обеспечивать у леммингов необходимый уровень зрелости и лабильности адаптивных реакций у плодов и новорожденных.
Это подтверждалось данными о незначительной эмбриональной смертности на стадиях с низкой плотностью популяции (Ткачев и др., 1987). Безусловным достижением работ популяционных экологов второй половины и конца прошлого века было продуктивное соединение знаний о вызываемых стрессом, в том числе СТМ, нарушениях развития (созревания) организма с наблюдаемыми особенностями динамики численности и демографическими изменениями в популяциях.
Эти работы имели отчетливую эволюционную направленность и рассматривали стресс как популяционную адаптацию, обеспечивающую настройку популяционной системы к модальным условиям ее существования (Шилов, 1984). Вместе с тем трактовки роли стресса как фактора, регулирующего динамические процессы в популяциях, выглядели скорее как гипотезы, нежели имели под собой количественные оценки приспособленности особей (выживаемость, репродуктивный успех), испытавших на себе стрессовые нагрузки разной модальности. Кроме того, наблюдения в природе не давали возможности судить о самостоятельной роли последствий стресса в раннем онтогенезе в регуляции популяций вне связи с общими, легко наблюдаемыми эффектами хронического стресса, ведущими к подавлению репродукции, иммунной системы, истощению, нарушению пищеварения и т.д. Новый подход обозначился в самом конце прошлого – начале нынешнего веков, когда начинают использоваться индивидуально ориентированные наблюдения и эксперименты в природе или в условиях, моделирующих природные ситуации. Важную роль в становлении нового подхода сыграла теория оптимизации распределения репродуктивного вклада в потомство (sex allocation theory; Charnov, 1982).
2. Оптимизация распределения репродуктивного вклада в потомство и пренатальный стресс
У ряда видов наземных позвоночных животных сильные, здоровые матери рождают больше сыновей в отличие от матерей со слабым или нарушенным здоровьем, у которых соотношение полов в выводках смещено в сторону преобладания дочерей. Объяснением этого явления может служить модель, предложенная Триверсом и Уиллардом (Trivers, Willard, 1973) как составная часть более общей теории оптимизации соотношения полов через распределение репродуктивного вклада в потомство (sex allocation theory; Charnov, 1982; West, 2009). Основная идея гипотезы Триверса и Уилларда состоит в том, что при плохих
условиях выгоднее вкладывать в дочерей, поскольку шанс размножения самки даже в пессимальных условиях достаточно высок, а “плохие” сыновья будут неконкурентоспособными и вероятность их размножения низка. Напротив, при хороших условиях существования выгоднее вкладывать в сыновей, поскольку здоровые сыновья имеют большую репродуктивную ценность, чем здоровые дочери (Trivers, Willard, 1973; Frank, 1990). Существует довольно много свидетельств того, что стрессовая среда и соответственно СТМ (пренатальный стресс) смещает соотношение полов в сторону преобладания самок.
Так, яйцекладущие самки ящерицы (Uta stansburiana), получавшие экзогенные глюкокортикоиды (кортикостерон) из имплантов в силиконовой оболочке, формировали более богатые энергоресурсами яйца, из которых вылуплялись относительно более крупные самки, однако это не касалось самцов (Sinervo, DeNardo, 1996). У европейского скворца инъекции кортикостерона в свежеотложенные яйца приводили к вылуплению самок более высокого качества по сравнению с контролем, но это не касалось вылупившихся самцов. Самцы из инъецированных кортикостероном яиц появлялись на свет более мелкими, медленнее росли и отличались повышенной смертностью в раннем возрасте, несмотря на то, что в норме у европейского скворца самцы крупнее самок (Love, Williams, 2008a).
У потомков самок крыс, подвергавшихся в период беременности социогенному стрессу (чрезмерная плотность группы), наблюдалась феминизация поведения сыновей, сокращение у них ано-генитального расстояния по сравнению с контрольной группой, но у самок видимые изменения поведения на протяжении эстрального цикла отсутствовали (Dahlöf et al., 1977). У беличьих саймири, самки которых в период беременности испытывали пренатальный стресс (помещение в чужую группу), у сыновей наблюдались признаки иммуносупрессии. Концентрация иммуноглобулина IgG у сыновей падала, в то время как у дочерей, напротив, увеличивалась (Coe et al., 2000).
Признаки инфантилизации сыновей и маскулинизации дочерей были подробно исследованы у морских свинок (Kaiser, Sachser, 2005; Sachser et al., 2011; подробнее см. ниже). Связь пренатального стресса как высокой, так и средней модальности со смещенным в сторону преобладания самок соотношением полов при рождении была показана в популяциях людей (Peterka et al., 2004; Obel et al., 2007). Не исключено, что более частое рождение девочек в некоторых странах является следствием стрессов, испытываемых матерями в период беременности (Obel et al., 2007). Результаты приведенных выше исследований (перечень примеров может быть продолжен) поддерживают гипотезу Триверса и Уилларда (Trivers, Willard, 1973) и свидетельствуют в пользу приспособительного значения смещения соотношения полов у потомков, матери которых испытывали стресс.

Беличий саймири Saimiri sciureus
Существует, однако, немало исследований, в которых интерпретировать последствия пренатального стресса как благоприятствующие дочерям и негативные в отношении сыновей, либо как положительные в отношении потомков одного пола и отрицательные – другого пола, в чистом виде не удается (Allen, Haggett, 1977; Harvey, Chevins, 1984, 1985, 1987; Zielinski et al., 1991; MarchlewskaKoj et al., 2003a, b; Kaiser, Sachser, 2005, и др.). Эти исследования проводились в условиях лаборатории на немногих модельных видах млекопитающих. Результаты этих работ, даже если и демонстрируют в той или иной степени признаки феминизации или инфантилизации сыновей и маскулинизации дочерей (либо отсутствие того или другого), в основном свидетельствуют о нарушении нормального полового развития, о замедлении темпов полового созревания. В этих исследованиях эффектов пренатального стресса внимание, как правило, сосредоточено на программировании мозга, функциональных связей ГГНС или поведения (Weinstock, 2005, 2007, 2008; Glover et al., 2010; McEwen et al., 2012).
Сравниваемые признаки потомков в опыте и контроле в этих работах могут иметь весьма отдаленное отношение к реальным индикаторам приспособленности самцов и самок, рожденных матерями, испытавшими пренатальный стресс. Кроме того, возможны и иные причины нарушения эмбрионального соотношения полов. У водяной полевки (Arvicola terrestris) преобладание в помете самцов, а не самок может быть связано с низкой энергообеспеченностью матерей и, как следствие, повышенной эмбриональной смертностью дочерей (Назарова, Евсиков, 2000; Назарова, 2008). Авторы рассматривают это явление как адаптивное, поскольку снижается вероятность рождения низкокачественных самок матерями низкого качества (качества, определяющие успешность выкармливания потомства, наследуются дочерьми: h2 = 0.64).
3. Эпигенетическое программирование морфофункциональных и поведенческих качеств потомства у рыб
3.1. Рыбы коралловых рифов, социальная конкуренция и трэйдоффы между выживаемостью и смертностью
Рыбы тропических рифов обитают в условиях высокого разнообразия видов и высокой плотности населения в период размножения и поэтому представляют собой удобную модельную группу для изучения внутри- и межвидовых, обусловленных социальной средой влияний материнских глюкокортикоидов (СТМ) на фенотипическую изменчивость и приспособленность потомства (McCormick, 1998). У тропической рыбы-ласточки (Pomacentrus amboinensis) из сем. Pomacentridae самцы охраняют придонные гнезда, в которые откладывают икру находящиеся поблизости самки. Между самками существуют иерархические отношения. Большая часть отложенной икры принадлежит доминирующей самке. Часть самок икру не откладывает. Однако общая плотность самок, взаимодействующих с размножающимися самцами, положительно влияет на уровень кортизола в организме последних. Повышенный уровень кортизола материнского организма обусловливает появление из икры мелких личинок (McCormick, 1998, 2006); при этом объем желтка в желточном мешке остается неизменным (Gagliano, McCormick, 2009). Во время пелагической стадии, которую проходят личинки (между 15 и 23-м днями), наблюдается интенсивная, зависящая от размера особи смертность, причем наибольшей выживаемостью характеризуются личинки мелкого размера, но с относительно большими желточными мешками (Gagliano et al., 2007).

Pomacentrus amboinensis
При высокой локальной плотности рыб выселение из мест рождения имеет несомненные выгоды, но требует в целом более выносливой личинки, т. е. большого резерва желтка, дающего возможность длительного пребывания в состоянии пелагической стадии. Таким образом, в противоположность мнению о том, что “быть большим всегда выгодно”, опосредованная плотностью индуцированная глюкокортикоидами редукция размера вышедшей из икринки личинки адаптивна, поскольку сочетается с относительно большими резервами желтка. И наоборот, больший размер, высокие темпы роста личинки на начальной стадии формирования сообщества увеличивают выживаемость (Holmes, McCormick, 2006; Gagliano et al., 2007), вероятно, за счет снижения уязвимости таких мальков для хищника (gape limited predation).
Участки с низкой плотностью рыб и редкими социальными взаимодействиями населены самками с низкой глюкокортикоидной реакцией. Эти самки производят мальков большего размера (McCormick, 1999, 2006), для которых сокращение дистанции расселения представляется выгодным. Таким образом, у рыбы-ласточки действует плотностнозависимый механизм, оптимизирующий расселение и пополнение локальной популяции (сообщества) наиболее конкурентоспособными рыбами. Эти же работы содержат результаты, указывающие на то, что для реалистичной объективной оценки адаптивной ценности индуцированных материнским эффектом изменений в размерных и других характеристиках потомков, приспособленность должна оцениваться на протяжении всей жизни особи. Влияния пренатальной среды, которая индуцировала фенотипические изменения у потомков, безусловно, велико, но не только пренатальная среда определяет индивидуальную приспособленность.
3.2. Трехиглая колюшка и хищники – формирование адаптивного профиля поведения мальков путем модификации развития икры под влиянием материнского стресса
В исследовании Гизинга с соавторами (Giesing et al., 2011), выполненном на трехиглой колюшке (Gasterosteus aculeatus), имитировалось нападение хищника – щуки (Esox lucius) – на рыбок экспериментальной группы один раз в день в разное время суток. Оказалось, что подвергавшиеся угрозе нападения самки откладывали более крупную икру с повышенным содержанием кортизола. Такие икринки потребляли больше кислорода сразу после оплодотворения по сравнению с контрольными (в контроле самки, отметавшие икру, атакам хищника на стадии формирования и созревания икры не подвергались). Мальки, вышедшие из икры, отложенной самками, подвергавшимися нападению щуки-модели, предпочитали держаться более плотной группой.
Такое поведение экспериментальной группы мальков адаптивно, поскольку обеспечивает лучшую защиту молоди от хищников (Magguran, 1990; Queiroz, Magurran, 2005). При этом материнский эффект не сказывался на соматическом росте мальков (Giesing et al., 2011). Реакция самок колюшки на присутствие хищника также влияла на способность потомства к обучению (Roche et al., 2012). Из четырех гормонов материнского происхождения в икре колюшки уровни прогестерона, тестостерона и эстрадиола резко снижались после оплодотворения икры и оставались на низком уровне. Однако уровень кортизола снова поднимался на восьмой день после оплодотворения (Paitz et al., 2015).
Изменение уровня стероидов было обусловлено эмбриональными процессами, менявшими первичную эндокринную среду эмбриона. Эндокринные изменения эмбриональных стадий подтвердили способность материнских стероидов влиять на развитие организма потомков. Проведенные исследования показали, что воздействие экологически значимого стрессора на самку трехиглой колюшки во время формирования икры имеет далеко идущие последствия для потомства. Некоторые из этих эффектов могли быть опосредованы воздействием кортизола, уровень которого повышался в организме самки под воздействием хищника.
Вопрос, в какой мере глюкокортикоидный профиль материнского организма определяет глюкокортикоидный профиль подросшего потомства, исследовали в специально поставленном эксперименте. В этом случае охоту хищника моделировали сначала применительно к самкам-матерям, а потом уже применительно к их подросшей молоди (Mommer, Bell, 2013). Хотя потомство матерей, подвергавшихся воздействию хищника, отличалось по глюкокортикоидному ответу от потомства контрольных матерей в ситуации предъявления хищника потомству, оказалось, что глюкокортикоидная реакция потомков на хищника в значительно большей степени определялась полом и социальной средой.
Среди потомства самки имели более высокий уровень кортизола, чем самцы, а рыбы в группе имели более низкий уровень кортизола, чем рыбы, которые были изолированы от конспецификов. В совокупности результаты этих исследований однозначно свидетельствуют в пользу того, что самки трехиглой колюшки могут адаптивно влиять на развитие, рост и поведение потомства, обеспечивая соответствие его качеств вызовам среды в будущем.
Однако необходимо иметь ввиду, что это соответствие возможно лишь в случае совпадения текущих (в период вынашивания икры) и будущих условий, в которых будет жить следующее поколение. Если особи трехиглой колюшки, матери которых подвергались атакам хищника, сталкивались с реальным хищником – живой щукой – один на один, то они чаще становились жертвами щуки по сравнению с потомками матерей, не имевших опыта общения с хищником (McGhee et al., 2012).
Кроме того, выживаемость молодых колюшек зависела от их размера. Чем более крупными были колюшки, рожденные матерями, подвергавшимися воздействию хищника, тем более уязвимыми при столкновении с щукой один на один они оказались по сравнению с потомками матерей, не имевших опыта общения с хищником (McGhee et al., 2012). Таким образом, опыт встречи с хищником может иметь долгосрочные последствия для потомства, но их адаптивность зависит от контекста.
3.3. Родительская забота у рыб и социальное поведение потомков: программирование нейроэндокринной оси стресса у цихлид Neolamprologus pulcher
Регуляция экспрессии глюкокортикоидных рецепторов в раннем онтогенезе – возможный механизм вариации социальной конкурентоспособности (social competence). Группа исследователей под руководством Барбары Таборски (Nуman et al., 2018) проверила эту гипотезу на социальной цихлидовой рыбе Neolamprologus pulcher. Мальки этой цихлиды подращивались с участием и без участия родителей.

«Принцесса Бурунди» Neolamprologus pulcher
Молекулярный механизм, транслирующий ранний социальный опыт на зрелые стадии жизненного цикла, у рыб в общих чертах гомологичен таковому у млекопитающих (Steenbergen et al., 2011). Рыбки N. pulcher, воспитывавшиеся с родителями, имели более выраженную экспрессию гена глюкокортикоидного рецептора (ГР1) в переднем мозге. При этом уровни экспрессии гена транскрипционного фактора реакции раннего роста 1 (eGR1 – transcription factor of early growth response 1) были связаны с экспрессией ГР1 в переднем мозге и гипоталамусе. (Ген транскрипционного фактора eGR1 является маркером нейрональной активности и пластичности и сам кодирует транскрипционный фактор, который, как предполагается, активирует в дальнейшем гены кортикостероидной оси стресса в дорзолатеральной области переднего мозга рыб (Desjardins, Fernald, 2010)).
Блокирование глюкокортикоидных рецепторов антагонистом мифепристоном (RU486) приводило к тому, что выращенные родителями рыбки демонстрировали более социально адекватное, субмисcивное поведение в случаях, если оказывались на территории более крупных конспецификов. Эти рыбки реже бывали атакованы хозяевами территории, а в конечном счете оказывались более успешными в овладении территорией. Результаты этого исследования показали, что ранняя социальная среда программирует нейроэндокринную ось стресса у рыб (Taborsky et al., 2013; Nyman et al., 2017) подобно тому, как это происходит у млекопитающих (Wingfield, Sapolsky, 2003; Kaiser, Sachser 2005; Weinstock, 2008; Steenbergen et al., 2011; McEwen, 2012) и птиц (Banerjee et al., 2012; Zimmer, Spencer, 2014).
4. Плотность населения ящериц и филопатрия
Исследования ящериц, связавшие повышенную гормональную активность ГГНС размножающихся самок с фенотипическими эффектами, наблюдаемыми у молодняка, внесли значительный вклад в формирование представлений о возможной адаптивной роли последствий СТМ (Dufty et al., 2002). В исследовании, выполненном Синерво и ДеНардо (Sinervo, DeNardo, 1996), самкам яйцекладущей ящерицы (Uta stansburiana) вводили экзогенные глюкокортикоиды (кортикостерон) с помощью имплантов в силиконовой оболочке. Это не влияло на размер кладки, однако вело к формированию более богатых энергоресурсами яиц и вылуплению более крупных самок, но не самцов. При этом индуцированное имплантированным кортикостероном изменение текущих репродуктивных затрат вело к снижению выживаемости самок-матерей. Направление эффекта зависело от плотности населения ящериц в текущем году (Sinervo, DeNardo, 1996).
Поскольку концентрация кортикостерона в плазме крови размножавшихся самок ящериц была положительно связана с плотностью населения, было высказано предположение о том, что увеличение размеров вылупившихся молоденьких самок адаптивно, поскольку способствует выживанию молодняка в условиях жесткой конкуренции со взрослыми ящерицами (Comendant et al., 2003). В другом исследовании у живородящей обыкновенной ящерицы (Lacerta vivipara) наблюдался отрицательный гормонально обусловленный материнский эффект в отношении молодых самцов; самцы рождались более мелкими, имели худшие экстерьерные признаки и медленнее росли. При этом у них наблюдалась более высокая выживаемость (Meylan, Clobert, 2005; Coté et al., 2006). Кроме того, у потомков, рожденных старыми ящерицами, отличавшимися худшими экстерьерными качествами и повышенным уровнем глюкокортикоидов, наблюдалась склонность к филопатрии (Fraipont et al., 2000; Meylan et al., 2002).
Авторы предположили, что хронический подъем уровня глюкокортикоидов в организме самки во время беременности служит сигналом, свидетельствующим о низкой вероятности выживания матери после появления на свет молодых ящериц. Это в конечном счете обеспечивает меньшую напряженность конкуренции между родителями и потомством (Meylan, Clobert, 2005). Таким образом, уровень глюкокортикоидов в организме матери может влиять на динамические процессы как в локальной популяции, так и в мета-популяции в целом, влияя на формирование пополнения и другие процессы жизненного цикла (Meylan et al., 2012).
5. Европейский скворец и гипотеза “комплементарности материнского качества и требований потомства” (Matternal Match Hypothesis)
Интересный цикл исследований был проведен учеными из Канады и США в естественной популяции европейских скворцов (Sturnus vulgaris) в Канаде (Love et al., 2005; Love, Williams 2008a, b; Love et al., 2013). Эксперимент в природе был спланирован таким образом, что позволял одновременно манипулировать фенотипом потомства через стресс организма матери и условия жизни, с которыми птенцам предстоит столкнуться после появления на свет.

Поющий сковрец
Известно, что эмбриональные эффекты избыточных глюкокортикоидов материнского организма, как правило, снижают качество потомства. Однако, согласно идее авторов исследования, эффект может повышать совокупную приспособленность самки в случае комплементарности качества материнского организма и требований появившегося на свет потомства. Основной вопрос формулировался следующим образом: при каких условиях и в какой мере индуцированные материнским стрессом особенности потомства у европейского скворца могут быть полезными их матерям?
Кортикостерон инъецировали прямо в желток свежеотложенных яиц, моделируя таким образом эффект избыточных глюкокортикоидов (ГК) материнского организма. Помимо этого экспериментально меняли материнскую способность эффективно выкармливать птенцов. Предполагалось, что самки низкого качества, с высоким фоновым уровнем глюкокортикоидов, хуже выкармливают птенцов по сравнению с самками высокого качества, у которых фоновая “стрессированность” ниже. Отрицательная связь между родительскими качествами матери и базальным уровнем ГК материнского организма была показана ранее (Love et al., 2005; Love, Williams, 2008b).
Кроме того, уровень ГК материнского организма был положительно связан с уровнем ГК в яичном желтке (Love et al., 2005). Разнокачественность самок моделировали путем обрезания некоторых перьев в оперении на стадии выкармливания птенцов (Love, Williams, 2008a). В эксперименте были использованы четыре группы птиц:
1) птенцы, вылупившиеся из яиц, инъецированных раствором кортикостерона в кунжутовом масле, и их матери с обрезанными перьями,
2) птенцы, вылупившиеся из яиц, инъецированных кортикостероном, и их матери с необрезанными перьями,
3) птенцы, вылупившиеся из яиц, инъецированных чистым кунжутовым маслом, и матери с обрезанными перьями и
4) контрольные, без инъекции, птенцы с их полноценными матерями.
Далее исследовали, как ГК-индуцированное качество птенцов и качество их матерей влияют на плодовитость и выживаемость матерей в будущем. Экспериментально модулированный уровень желточных ГК вызывал связанный с полом трейдофф между количеством и качеством потомства. Повышение уровня кортикостерона в яйцах обусловливало более высокое качество дочерей. Напротив, самцы европейского скворца, в норме более крупные и быстро растущие по сравнению с самками, вылуплялись более мелкими, медленнее росли и отличались повышенной смертностью в раннем возрасте. Однако когда самцов, подвергшихся воздействию материнского стресса (инъекция кортикостерона в яйцо), выкармливали матери низкого качества (с обрезанными перьями), те из самцов, кто выжил, превзошли контрольных самцов по ряду фенотипических показателей-индикаторов приспособленности (Love, Williams 2008a; Chin et al., 2009). Хотя матери, у которых потомки испытали воздействие материнского стресса, выращивали меньшее число детей в текущем цикле размножения, соответствие качества матери требованиям потомства приносило матерям пользу в будущем. Самки в конечном счете извлекали выгоду из совокупного успеха размножения вследствие трейдоффов между текущими репродуктивными затратами матери и 1) ее будущим состоянием, 2) ее будущим репродуктивным вкладом, 3) выживаемостью.
Конкретно, самки низкого качества, как бы испытывавшие хроническую стрессовую нагрузку (оценивалась косвенно через ухудшение эффективности фуражировки и кормления птенцов в результате обрезания перьев) и вырастившие потомство, испытавшее воздействие экзогенных глюкокортикоидов,
1) находились в лучшем состоянии по экстерьерным показателям во время вторых выводков,
2) в будущем производили большее число потомков лучшего качества и
3) отличались более высокой выживаемостью в последующие годы (межгодовые сравнения; Love, Williams, 2008a).
Таким образом, анализ соответствия фенотипических характеристик потомства качеству материнской заботы продемонстрировал увеличение материнской совокупной приспособленности, несмотря на то, что некоторые из ответных реакций потомков в краткосрочной перспективе (снижение размера и темпов роста) выглядели неадаптивными (Love, Williams, 2008a).
Европейский скворец – не единственный среди воробьиных птиц пример адаптивных фенотипических изменений у потомков, матери которых подвергались стрессирующим воздействиям. В эксперименте в природе с большими синицами (Parus major), самкам которых предъявляли хищника, птенцы, выращенные другими родителями (не подвергавшимися риску нападения), отличались от контрольных кросс-фостеров меньшими размерами и весом. Интересно, что у первых быстрее развивались крылья, что, с точки зрения авторов исследования, возможно обеспечивало им больше шансов для выживания (Coslovsky, Richner, 2011).
Концентрация тестостерона в яйцах от самок, испытавших риск нападения хищника, была более низкой по сравнению с контролем. Различия по андростенону и прогестерону в желтке отсутствовали (Coslovsky et al., 2012). Однако известно, что именно тестостерон желтка влияет на темп роста и развития. Пониженный уровень тестостерона в желтке мог быть обусловлен гормонами стресса, концентрацию которых в данном исследовании не измеряли.
Опосредованное стрессом изменение уровня тестостерона в желтке могло модулировать дифференцированное развитие морфологических структур.
6. Влияние нестабильной социальной среды во время беременности на развитие потомков у морских свинок
Морская свинка была одомашнена в Южной Америке где-то между 6000 и 3000 лет назад. Несмотря на это, поведение домашней морской свинки (Cavia aperea f. porcellus) в общих чертах сходно с поведением дикого предка, C. аperea. Различия касаются в основном частоты проявления паттернов и порогов включения тех или иных форм поведения (Künzl, Sachser, 1997; Sachser, 1998). Социальные взаимодействия у домашних морских свинок отличаются большей дружелюбностью, а система иерархических связей при содержании группой более мягкая. При содержании в неволе самцы диких свинок крайне агрессивны по отношению к молодым созревающим самцам (вплоть до их гибели). Среди молодых самок складывается строгая линейная иерархия с доминированием старшей самки. Дикие морские свинки отличались в 5–7 раз более высоким уровнем адреналина и норадреналина в сыворотке крови в ответ на взятие в руки при сходном уровне кортизола (Künzl, Sachser, 1997).
Обширный цикл исследований влияния стресса в раннем онтогенезе на развитие мозга, формирование нейроэндокринных связей и поведения был выполнен группой немецких ученых на одомашненных морских свинках (Кaiser, Sachser, 2005, 2009; Sachser et al., 2011). В качестве стрессора в период раннего онтогенеза использовалась нестабильность социальной среды. Социально стабильные группы свинок состояли из пяти самок и одного самца. Социальная нестабильность в группе в период беременности и лактации обеспечивалась пересаживанием части самок из группы в группу (каждые три дня заменяли двух самок в группе). Было показано, что биосоциальные стимулы оказывают на морских свинок разнообразные нейроэндокринные эффекты (Hennessy, 1988; Sachser et al., 1994, 1998). В частности, нестабильная социальная среда в период беременности (пренатальный стресс) значимо влияла на развитие эндокринной системы, мозга и поведения потомства, причем это влияние было специфичным в отношении пола (Sachser, Kaiser, 1996; Kaiser, Sachser, 1998, 2001; Kaiser et al., 2003a, b).
Дочери самок, которые во время беременности испытали влияние социальной нестабильности, отличались от контроля (беременность протекала в социально стабильной среде) ярко выраженными признаками “поведенческой маскулинизации”. Для них были характерны позы и движения ухаживания, характерные для полового поведения самца, а также игровое поведение в период взрослой жизни (Sachser, Kaiser, 1996; Kaiser, Sasher, 1998; Kaiser et al., 2003a). Поведенческая маскулинизация дочерей сопровождалась повышенным уровнем тестостерона и повышенной адреномедулярной активностью, о чем свидетельствовала повышенная активность тирозин гидроксилазы мозгового вещества надпочечников (Kaiser, Sachser, 1998). Дочери, матери которых подвергались в период беременности влиянию нестабильной социальной среды, демонстрировали типичную для самцов экспрессию рецепторов андрогенов в медиальной преоптической области мозга (MPOA) и дугообразном ядре (Nucleus arcuatus, ARC) гипоталамуса (Kaiser et al., 2003a). Кроме того, у дочерей, чьи матери имели опыт жизни в нестабильной социальной среде во время беременности, было обнаружено усиление рецептора андрогенов в CA1-зоне гиппокампа, а также рецептора эстрогена-α (ER-α) в MPOA, ARC и CA1 (Kaiser et al., 2003a).
В отличие от пренатально стрессированных самок самцы, матери которых жили в нестабильной социальной среде, демонстрировали особенности поведения, характерные для молодых самцов – тесные телесные контакты во время отдыха.
У пренатально стрессированных самцов эта особенность сохранялась до более взрослого возраста. Кроме того, типичное половое поведение самцов (позы и движения ухаживания) часто оказывалось интегрированным в игровое поведение (Kaiser, Sachser, 2001). Таким образом, поведение пренатально стрессированных самцов имело черты инфантильного. Поведенческая инфантилизация (behavioral infantilization, Kaiser, Sachser, 2001) была связана с отсроченным развитием адренокортикальной системы и пониженной активностью симпатической адреномедулярной системы (Kaiser, Sachser, 2001). Самцы, чьи матери имели опыт жизни в нестабильной социальной среде, имели пониженную экспрессию рецепторов андрогенов в MPOA, ARC гипоталамуса, а также ER-α в CA1-зоне гиппокампа (Kaiser et al., 2003b).
Возможность трактовки результатов этих исследований как адаптивных по мнению авторов обоснована тем, что популяционная плотность морских свинок в природе варьирует в широких пределах (Sachser, 1998), а социальная организации меняется в зависимости от плотности (Sachser, 1986). Предполагалось также, что высокая популяционная плотность сопряжена с социальной нестабильностью (Von Holst, 1998). Какое приспособительное значение может иметь маскулинизация дочерей? Авторы исследования дают следующий ответ (Кaiser, Sachser 2005; Kaiser, Sachser 2009; Sachser, Hennessy, Kaiser, 2011). Самка морской свинки, живущая в условиях высокой плотности в социально нестабильной среде, эпигенетически программирует развивающийся организм потомка, подготавливая его к жизни в условиях высокой плотности. Маскулинизация дочерей адаптивна в плотных поселениях, поскольку самки становятся более конкурентоспособными в условиях дефицита пищевых ресурсов и убежищ (Кaiser, Sachser, 2005). Однако максулинизация поведения одновременно ухудшает и будущие репродуктивные качества самки.
Таким образом, в условиях высокой плотности действует трейдофф между маскулинизированным фенотипом на эндокринном и поведенческом уровнях и платой за снижение репродуктивного успеха. В условиях же низкой популяционной плотности более выгодно инвестировать ресурсы времени и энергии в репродуктивное усилие, нежели поддерживать более конкурентоспособный в дефицитной по обеспеченности пищей среде маскулинизированный фенотип.
Для сыновей же, рожденных испытавшими пренатальный стресс самками, в условиях высокой плотности более выгодно иметь менее выраженные признаки самца, в особенности в период, сопряженный с половым созреванием. Начальный период половой зрелости у самцов сочетается с напряженными социальными взаимодействиями, с частыми травмами при изменении позиций в системе социальных связей. В это время молодые самцы у морских свинок подвергаются жесткому агрессивному давлению со стороны взрослых самцов в борьбе за возможность размножения и вынуждены эмигрировать за пределы социума. В норме морские свинки созревают в возрасте 2–3 мес (Webster, Young, 1951).
С этим возрастом связано резкое увеличение агрессивных контактов у молодых самцов в присутствии рецептивной самки (Sachser, 1986). При низкой плотности самки максимизируют собственную приспособленность, пренатально программируя сыновей к раннему созреванию (раннее прекращение периода инфантильности). Напротив, в условиях повышенной популяционной плотности при наличии структурированной по возрастам социальной среды замедление созревания сыновей выгодно как матерям, так и их сыновьям. В этих условиях более 85% детенышей в экспериментальных разновозрастных группах домашних морских свинок принадлежат высокоранговым самцам (Sachser, 1998). При этом молодые самцы не созревают до возраста 7 мес и избегают агонистических взаимодействий в молодом возрасте (Sachser, 1993). В противном случае их репродуктивный успех был бы ниже того, который они смогут иметь в будущем. Половое созревание и социальная зрелость у морских свинок не вполне совпадают во времени. В результате поведенчески инфантильные, хотя уже и созревшие самцы, реже подвергаются атакам со стороны самца-доминанта (Sachser, Renninger, 1992).
Таким образом, пренатальное программирование сыновей матерями, которое выражается в меньшей экспрессии признаков самца и отсроченном созревании, обеспечивает им оптимальную репродуктивную стратегию в условиях высокой плотности населения.
Резюмируя итоги приведенных выше экспериментальных исследований одомашненных морских свинок, нельзя не отметить умозрительность и излишний адаптационизм трактовок авторами результатов. Так, предполагается, что самки при высокой плотности более конкурентоспособны в соревновании за ресурсы пищи, что адаптивно; они же при высокой плотности меньше вкладывают в потомков, которые обречены на гибель; таким образом, самки экономят ресурсы для будущего размножения – тоже адаптивно. Однако сам по себе отказ от размножения в стрессовой среде мало о чем говорит без демонстрации того, что совокупная приспособленность таких самок увеличивается.
7. Американский заяц-беляк (Lepus americanus) и 10-летний цикл в системе “хищник–жертва”
Среди свободноживущих млекопитающих яркий пример влияния материнского стресса (СТМ) на параметры физиологии и поведения жертвы, находящейся под влиянием хищника, численность которого меняется во времени, предоставляет американский заяц-беляк (Sheriff et al., 2009, 2010, 2011). Для популяций зайцев характерны регулярные циклические колебания численности с интервалом от 8 до10 лет между пиками (Krebs et al., 1995). Численность популяции хищника (в основном это американская рысь) отстает от численности популяции жертвы на 1–2 года. В периоды падения численности зайцев большинство из них гибнет от хищников. После окончания фазы падения численности популяция зайцев продолжает оставаться в состоянии глубокой депрессии еще от 2 до 5 лет (восстановление не начинается), несмотря на низкую численность хищников и хорошую обеспеченность зайцев кормом (Krebs et al., 1995). Максимально успешное размножение зайцев происходит на начальной стадии восстановления их численности, когда численность хищников минимальна; успешность размножения зайцев прогрессивно снижается в течение фазы падения их численности, достигая состояния минимума при максимальной численности хищников. Успех размножения зайцев остается минимальным до конца фазы депрессии (окончание фазы низкой численности; Krebs et al., 1995).
Во время фазы спада численности вызовы среды (направления воздействия факторов) в отношении матерей и их потомков совпадают. Высокий риск нападения хищника обусловливает повышенный уровень стрессированности матерей, о чем свидетельствует высокий уровень глюкокортикоидов в материнском организме (Sheriff et al., 2011). С высоким материнским стрессом связано уменьшение размера выводков и рождение зайцами более мелких детенышей (Sheriff et al., 2009).
Эта связь прослежена как в природе, так и в эксперименте. Более того, экспериментально показано, что пренатально-стрессированные зайчата отличаются более высоким уровнем глюкокортикоидов и более интенсивной реакцией на стрессоры по сравнению с зайчатами, не подвергавшимися влиянию СТМ. Под влиянием СТМ зайчата становятся более пугливыми и настороженными. Такое изменение поведения зайчат способствует увеличению их выживаемости (Sheriff, Love, 2013). Поведенческие особенности сохраняются и во взрослом возрасте (Sheriff et al., 2010). Одновременно более низкая успешность текущего размножения самок коррелирует в условиях высокой вероятности нападения хищника с их лучшей выживаемостью в основном за счет сокращения времени на поиск и добывание корма. При этом продолжительность периода активности самок сокращается за счет времени, когда атака хищника наиболее вероятна.

Американский беляк
В фазе роста численности популяции зайца направления воздействия факторов среды на зайчих и на их потомство также совпадают. Эти вызовы среды определяются низким риском быть убитым хищниками (хищников еще мало) и высоким обилием пищи (численность зайцев еще низка). В этих условиях уровень глюкокортикоидов у беременных зайчих низкий. Они рождают крупных зайчат с низкой стресс-реакцией на хищника. В результате в этой фазе движения численности приспособленность зайчих высокая. Принципиально иная ситуация складывается в начале фазы депрессии. В этот период движения численности популяции американского зайца-беляка наблюдается несовпадение (mismatch) вызовов среды в отношении беременных зайчих и их потомков (матери испытывали повышенный стресс от хищников в конце фазы падения численности, а их потомки уже испытывают низкий пресс хищников в фазе депрессии). Зайчихи производят на свет выводки меньшего размера и более мелких детенышей с повышенной стресс-реактивностью. Эта ситуация не адаптивна. Комбинация эффектов вследствие несоответствия вызовов, предъявляемых средой матерям в период беременности, и вызовов среды обитания их потомства является, по мнению авторов исследования, основной причиной задержек восстановления численности зайцев, т.е. затяжных, до 5 лет, периодов депрессии, когда численность хищников уже низка, а ресурсы пищи в избытке (Sheriff, Love, 2013). Этот лаг, удлиняющий цикл численности, характерен не только для американских зайцев-беляков, но и для популяций полевок (Boonstra et al., 1998). Индуцированная СТМ неадаптивная фенотипическая изменчивость в начальный период депрессии численности может быть объяснением затяжных периодов депрессии численности и у других видов млекопитающих.
Таким образом, индуцированные СТМ фенотипические особенности потомства, проанализированные с учетом средовых контекстов, свидетельствуют в пользу адаптивных преимуществ СТМ фенотипов, однако лишь в случае, когда вызовы среды в отношении матерей и потомков совпадают, и наоборот – о неадаптивности эффектов материнского стресса, если такое соответствие отсутствует (Sheriff, Love, 2013).
8. Последствия пренатального стресса у людей: патологии или адаптации
Стресс, испытываемый матерью во время беременности, оказывает разнообразные долговременные эффекты на детей, которые в современных условиях жизни проявляются как разного рода нарушения (в развитии мозга, психофизиологии, поведении и т.д.) (Talge et al., 2007; Beydoun, Saftlas, 2008; Wadhwa et al., 2009, и др.).
Различные психопатологии у таких детей включают избыточную тревожность (increased anxiety; Van den Bergh, Marcoen, 2004; Bergman et al., 2007; Bruijn et al., 2009, и др.), дефицит внимания или гиперактивность (attention deficit/hyperactivity disorder (ADHD): O’Connor et al., 2002a; Obel et al., 2003a; Rodriguez, Bohlin, 2005; Van Den Bergh, Marcoen, 2004), кондуктивные расстройства, антисоциальные проявления, включающие повышенную агрессивность (conduct disorders: O’Connor et al., 2002b; Van den Bergh, Marcoen, 2004; Bruijn et al., 2009; Rice et al., 2010); ухудшение когнитивных способностей (Bergman et al., 2007; Laplante et al., 2008; Mennes et al., 2006, и др.), нарушенное кросс-доминирование правой и левой рук (more mixed handedness: Obel et al., 2003b; Glover et al., 2004; Rodrigues, Waldenstrom, 2008). Кроме того, имеют место различные нарушения функционирования ГГНС (O’Connor et al., 2005; Yehuda et al., 2005; Huizink et al., 2008; Van Den Bergh et al., 2008; Entringer et al., 2009).
Отмечено более редкое рождение сыновей, что является, по-видимому, следствием перераспределения репродуктивного вклада в потомков в пользу дочерей как более перспективного пола в условиях стресса (Peterka et al., 2004; Obel et al., 2007). Не исключено, однако, что многочисленные изменения, которые имеют вид патологий в настоящем, в эволюционном масштабе времени могли иметь приспособительное значение. В биомедицинских исследованиях идея о том, что развитие плода должно происходить в направлении адаптации к среде, в которой ему придется существовать, встречается в ряде работ (Gluckman et al., 2005, 2007; Glover, 2011). Гловер (Glover, 2011) специально рассматривает проблему постнатальных психопатологий в эволюционной перспективе. Хотя некоторые из предлагаемых ею объяснений могут показаться тенденциозно адаптационистскими, в целом эволюционный подход автора заслуживает внимания, поскольку по-новому расставляет акценты в традиционных биомедицинских исследованиях. Гловер считает, что в контексте эволюционного подхода становится более понятным, почему синдром повышенной тревожности, но не депрессия у матерей в период беременности, имеет более выраженный эффект у детей, почему повышенная тревожность и симптомы ADHD – наиболее частые эффекты пренатальных стрессов, почему депрессивные состояния и повышенная тревожность у детей проявляются при разных модальностях пренатальных стрессовых состояний матери и т.д. (O’Connor et al., 2002b; Field et al., 2003).
Очевидно, эпигенетическое программирование организма ребенка организмом матери в период эмбриогенеза (fetal programming, FP) идет различным образом в зависимости не только от стадии беременности, в которой организм матери испытывал стресс (соответствие времени формирования структур и функциональных связей в развивающемся организме), но и от природы материнского стресса. Тщательное изучение условий и FP возможно сможет помочь объяснить, почему некоторые психопатологии развития персистируют в популяциях человека. Не исключено, что в определенных обстоятельствах эволюционного прошлого было выгодным иметь в популяции долю индивидов с повышенной тревожностью, способных быстро переключать внимание, менять схемы поведения или быть более агрессивными. Так, ADHD синдром при рассмотрении его в контексте эволюционного прошлого человечества мог означать способность быстро перестраивать внимание в насыщенной опасностями среде, которая была стрессовой и для матери, и для ее потомков. Повышенная восприимчивость к сигналам слабой модальности, к новым звукам предупреждала о приближении хищника или врага. ADHD синдром означал повышенную готовность к исследованию и освоению новой среды, что тоже было важным этапом прогрессивной эволюции человека (Glover, 2011).
Гиперактивность и импульсивность означают непредсказуемость поведения. В изменчивой среде такое поведение отдельных индивидов в составе родового социума могло быть полезным (Williams, Taior, 2006). То же относится к синдрому повышенной тревожности в результате пренатального стресса. Повышенная тревожность (anxiety) в эволюционной истории человека означала повышенную настороженность (vigilance) и быстроту поведенческой реакции в условиях постоянной опасности, а также возможность подготовиться к оптимальному реагированию на стрессор средней интенсивности (Marks, Nesse, 1994). Кондуктивные расстройства могли означать повышенную готовность к освоению новой среды, более легкому (эмоционально) преодолению опасностей, избеганию хищников; повышенная агрессивность могла означать повышенную конкурентоспособность. Пониженные когнитивные способности ассоциированы с рассеянным вниманием, неспособностью концентрироваться на чем-то одном (distracted attention). Существуют свидетельства связи аутизма детей с пренатальным стрессом.
Но помимо нарушения способности к социализации аутизм – это еще и особый способ мышления. Нельзя исключить, что аутичные индивиды с синдромом средней модальности обладали когнитивными способностями, имевшими в эволюционном прошлом определенную выгоду. Наконец, измененная структура связей ГГНС означает изменение в порогах реагирования на стрессоры, которые очевидно были иными, чем в современном мире. Конечно, такого рода объяснения на самом деле объяснениями не являются. И если одни из них могут быть приняты в качестве рабочих гипотез, имеющих пусть косвенные, но свидетельства в их пользу (например, результаты изучения поведения настороженности (vigilance) у млекопитающих в природе; Quenette, 1990), то другие – чисто умозрительны, и объяснений, почему аутизм мог быть полезен в прошлом, можно придумать немало. Основная проблема состоит в том, что отсутствует или крайне ограничена возможность оценки “адаптивности” последствий пренатального стресса у детей в эксперименте. И все же работа в направлении поиска возможностей для экспериментальной проверки некоторых гипотез, как и поиск косвенных свидетельств значимости в эволюционном прошлом приобретенных в раннем онтогенезе нарушений поведения представляется оправданной. Древняя наследственная компонента стереотипов нашего эмоционального реагирования, разного рода фобий несомненно существует. Так, если рассматривать фобии взрослых людей, то очень непросто убедить человека не бояться змей, но гораздо труднее заставить его бояться автомобилей (Marks, Nesse, 1994).
Адаптивный потенциал стресса периода раннего онтогенеза
Проблема адаптивности последствий стресса раннего онтогенеза неразрывно связана с теорией “конфликта интересов родителей и потомков” (parent-offspring conflict theory: Trivers, 1974; Godfray, 1995; Parker et al., 2002), в рамках которой баланс интересов ближайших потомков и родителей решается в терминах затрат и выигрыша в ближней и отдаленной перспективе. По мнению Маршалла и Уллера (Marshall, Uller, 2007), в большинстве случаев конфликт интересов матери и потомков на интервалах времени, соизмеримых с продолжительностью жизни матери, решается в ее пользу вне зависимости от того, увеличивает или снижает материнский эффект выживаемость или успех размножения ее ближайших потомков.
Иными словами, материнский эффект в масштабе цикла текущего размножения может как увеличивать приспособленность потомков, т.е. быть адаптивным для них, так и уменьшать их приспособленность (неадаптивный эффект). Однако совокупная приспособленность матери в течение жизни может расти как за счет успешности будущего размножения матери, так и за счет будущего размножения отдельных выживших потомков. Оценка полезности материнского эффекта непосредственно для ближайшего потомства, т.е. индуцированной матерью адаптивной фенотипической изменчивости потомства, возможна лишь тогда, когда исследователь располагает данными об изменчивости окружающей среды во времени или в пространстве (если расселяющийся молодняк меняет условия своего обитания), когда условия окружающей среды для потомства предсказуемы относительно фенотипа матери и условий среды ее жизни, а затраты на передачу материнского сигнала или перераспределение ресурсов сравнительно низкие и, следовательно, напряженность конфликта интересов матери и интересов потомства не высока.
Применительно к обсуждаемой нами проблеме адаптивности последствий стресса раннего онтогенеза фенотипически проявляющиеся у потомков последствия ранних стрессов могут быть полезными для них в случае соответствия требованиям среды, с которыми потомки столкнутся в своей дальнейшей жизни. Это возможно лишь в том случае, если требования среды, в которой существовал организм матери в период формирования яйца или беременности и сразу после рождения выводка, совпадут с требованиями среды жизни ее потомков (Love et al., 2005; Love, Williams, 2008a; Sheriff, Love, 2013; Sheriff et al., 2009, 2010). В тех же случаях, когда условия среды обитания матери и ее потомства не совпадают, СТМ обусловливает неадаптивные фенотипические изменения у потомков, снижающие их приспособленность (Sheriff et al., 2009, 2010). При этом последствия СТМ, снижающие приспособленность потомства, могут быть выгодны для матери, поскольку оптимизируют ее затраты на текущее размножение, и в конечном счете могут повышать ее совокупную приспособленность (Love, Williams, 2008a). Правильная оценка адаптивной значимости СТМ возможна, таким образом, лишь в масштабе не отдельного цикла размножения, но всего жизненного цикла матери (integrated life history perspective, Sinervo, DeNardo, 1996).
Возникает вопрос, насколько реален адаптивный вектор в пре- и постнатальном программировании потомков, если рассматривать это явление в эволюционном масштабе времени? Может ли стресс, испытываемый в раннем онтогенезе, обеспечивать основу и задавать направление эволюционных преобразований? Множество исследований свидетельствует о росте фенотипической и генетической изменчивости при стрессе. Эта изменчивость служит материалом для формирования новых адаптаций (Шмальгаузен, 1968; Shapiro, 1992; Jablonka, Lamb, 1995; Hofman, Parsons, 1997, и др.). Стрессовая среда дестабилизирует и модифицирует интегрированную генетическую систему (Sharloo, 1991). Помимо мутагенных эффектов стресс, дестабилизируя систему онтогенеза, обусловливает фенотипическое проявление аккумулированной скрытой генетической изменчивости (Беляев, 1979, 1983; Беляев, Бородин, 1982; Bradshaw, Hardwick, 1989; Gibson, Wagnerr, 2000; Badyaev, 2005; Bijlsma, Loeschcke, 2005, и др.).
При стрессах периода раннего онтогенеза возникающие в результате пре- и раннего постнатального программирования фенотипические модификации, обусловленные эпигенетическими механизмами, также вносят существенный вклад в формирование фенотипического разнообразия, создавая дополнительные возможности для отбора. Благораря пластичности формирующихся структур и функций в процессе индивидуального развития организма пре- и постнатальное программирование дает возможность онтогенетической системе приспособиться к стрессовой среде. При этом индуцированная ранними стрессами изменчивость не случайна, но определенным образом канализирована (Badyaev, 2005). Эта канализированность вызванных стрессами раннего онтогенеза изменений задана онтогенетической системой и, сама являясь результатом предшествующей эволюции, служит одним из факторов быстрых эволюционных изменений в предсказуемо изменяющейся стрессовой среде (Badyaev, 2005).
В предсказуемо флуктуирующей среде она может служить объяснением существования альтернативных сценариев онтогенеза. Не исключено, что СТМ выступал, а возможно и сейчас выступает в качестве модификатора эволюционно обусловленной программы онтогенеза, запускаемой сигнальными факторами среды (развитие и созревание по быстрому или медленному сценариям) у сезонно размножающихся грызунов в зонах с умеренным и холодным климатом (Новиков, Мошкин, 2009). Эта же канализированность вызванной ранними стрессами вариабельности может обусловливать быстрые эволюционные изменения в случае однонаправленных изменений среды. Обусловленные эпигенетически, повторяющиеся модификации сохраняются в следующих поколениях, могут накапливаться в популяции и поддерживаемые отбором постепенно закрепляться на генетическом уровне. Так, трансгенерационные эффекты СТМ могут способствовать генетической ассимиляции стресс-индуцированных эффектов (Badyaev, 2009; Badyaev, Uller, 2009).
Вполне вероятно, что у видов с разным образом жизни в основе эволюционно детерминированных различий реагирования на стрессоры социальной природы лежали эпигенетические механизмы развития фенотипических проявлений стресса раннего онтогенеза. Чувствительность к стрессорам в постнатальной жизни потомков стрессированных матерей может быть как более высокой (низкий порог реагирования; Sherif, Love, 2013), так и пониженной (высокий порог реагирования; Дыгало и др., 1990).
Не исключено, что повышение порога чувствительности к стрессорам и модификация поведения в направлении к большей социальной терпимости в условиях повышенной плотности социальной среды в фазе роста и пика численности популяции давала возможность относительно безболезненно переносить агрессивные проявления со стороны других членов социума. Повышенная устойчивость к социогенным стрессорам у одиночных, некооперативно размножающихся видов млекопитающих могла оказаться первым шагом на пути формирования семейно-группового образа жизни (Попов, 2011). Известно, что у кооперативно размножающихся видов с наиболее устойчивыми системами доминирования-подчинения повышенный уровень глюкокортикоидов в большей степени свойственен особям, способным выдерживать высокие стрессовые нагрузки. Именно они становятся доминантами, определяющими структуру социальных связей в группе (Creel, 2001; Creel et al., 2013). Напротив, у одиночных, некооперативно размножающихся видов, более высокая стрессированность чаще встречается у субординантных особей. Случаи подавления репродуктивной функции стрессом описаны в основном у тех млекопитающих, для которых репродуктивная кооперация не характерна (Creel, 1996, 2001; Creel et al., 2013).
Заключение
Идея об обусловленной стрессом матери экономии затрат на текущее размножение не нова. В качестве выгоды для матери исследования классического периода популяционной экологии рассматривали возможность сохранения репродуктивного потенциала матери для ее размножения в будущем (Роговин, Мошкин, 2007). В качестве выгоды для потомства у быстро созревающих грызунов рассматривался отказ от размножения в текущем сезоне за счет развития по медленному сценарию, обеспечивающему лучшую подготовку к переживанию неблагоприятных зимних условий (Оленев, 1981, 2002; Новиков, Мошкин, 2009).
Возникает вопрос, что нового дают современные проверки адаптивности последствий стресса раннего онтогенеза по сравнению с трактовками классического периода популяционной экологии? Исследования раннего периода, если проводили в природе, то на кротких временных интервалах, и они имели характер констатации факта наличия связи между гипотетическим стресс-фактором и его последствиями. Результаты лабораторных экспериментов, в которых моделировали стрессовые ситуации, в том числе хронические, и оценивали их последствия (отказ от размножения, торможение развития и созревания
потомства, его повышенная смертность и пр.), экстраполировали на природные ситуации (Роговин, Мошкин, 2007). Действенным инструментом современных исследований адаптивности последствий стресса раннего онтогенеза у животных стал эксперимент в природе или в условиях, моделирующих природные (Sinervo, DeNardo, 1996; Lowe et al., 2005; Love, Williams, 2008a, b; Giesing et al., 2011, и др.).
Такие исследования остаются немногочисленными. Их инструментарий нельзя считать вполне сформированным. Однако уже сейчас обозначились общие особенности подхода.
1. Обязательный мониторинг судьбы поколений родителей и потомства (Sheriff, Love, 2013).
Долговременные эксперименты в природе дают возможность оценить приспособленность как матерей, испытывавших и не испытывавших повышенную стрессовую нагрузку, так и их потомков на фоне меняющихся условий жизни. Конечно, прямая оценка приспособленности через успех размножения (по числу потомков, достигших половой зрелости и вступивших в размножение) при работе с природными популяциями во многих случаях невозможна. Однако косвенные количественные оценки через выживаемость или успешность рекрутирования потомков в социум в сочетании с экспериментальными манипуляциями дают информацию, позволяющую делать статистически обоснованные заключения. Хорошо известно, что чувствительность к воздействию стрессоров меняется в течение жизненного цикла (Wingfield et al., 1998; Wingfield, Ramenofsky, 1999; Romero, 2002; Sherif, Love, 2013). Мониторинг последствий стресса раннего онтогенеза на больших интервалах времени важен еще и потому, что постнатальная среда жизни (качество материнской (родительской) заботы, опосредованной изменившимися условиями кормообеспеченности, климата, прессом хищников) может компенсировать эффекты пренатальной среды (Sherif, Love, 2013).
2. Обязательный учет условий жизни матери и потомства.
В отличие от лабораторного эксперимента эксперимент в природе не дает возможность стандартизировать условия опыта и контроля настолько, чтобы исключить их влияние на результат. Однако существует возможность оценить влияние факторов статистически, путем использования методов множественных сравнений, таких как общие линейные (GLM) и обобщенные (GLZ) модели. Последние позволяют анализировать как линейные, так и нелинейные эффе
Свежие комментарии