 Print
Print  PDF
PDF
Резюме. В продолжение темы копирования и «интуитивного понимания» действий другого, обслуживаемых зеркальными нейронами, см. соответствующий отрывок из книги их первооткрывателя. Фридрих Энгельс и Людвиг Нуаре были б ему особенно рады — среди прочего там показывается, как именно деятельность древних людей, в т.ч. трудовая, определила слова языка, возникшего несколько позже, и именно в плане членораздельности и акустической формы делений. Также как и идея синкинезии как модуса происхождения языка, опирающаяся на близкий корпус данных. Не менее важен вывод, следующий из этих данных, что «понимание Другого» связано с нервными, а не гуморальными механизмами, с копированием действий и извлечением из них концептов «типичного действия в типичной ситуации», а не влечением (или отвлечением), детерминированным гормональным состоянием, вроде обычных для гг.биологизаторов сентенций, что «чёрный» автоматически интерпретируется как «чужой», «опасный», а женщина — «сексуальный объект». Но этот механизм, обслуживаемый зеркальными нейронами, работает лишь в случае социального равенства, когда источник наблюдаемых действий воспринимается как «такой же как я». Если нет, он отключается, и нам кажется что «их» «нам никак не понять»: и самые опасные стереотипы против женщин, «цветных», других угнетённых групп работают именно на этом поле, утверждая что они «по природе другие«. Тогда внимательность к их поведению и копирование отключаются, как описано в книге — вместе с пониманием и эмпатией.

Джакомо Ризолатти
«Канонические и зеркальные нейроны
Содержание
Анализ функций зоны F5 показал нам, что большинство нейронов этой области коры разряжаются в процессе выполнения определенных действий, таких как хватание, удержание, манипулирование объектами, и что помимо этого часть из них отвечает на зрительные стимулы. Между моторными свойствами этих последних нейронов (например, специфическим типом захвата, который они кодируют) и их перцептивной избирательностью (к форме, размеру, ориентации объекта) существует тесная связь, которая определяет их важную роль в процессе преобразования зрительной информации о воспринимаемом объекте в моторные акты.
Эти нейроны были названы каноническими, потому что в течение долгого времени считалось, что премоторная кора может участвовать в зрительно-моторных преобразованиях. Однако, проведенные в начале 90-х годов эксперименты, в которых обезьян специально не учили выполнять задания, а позволяли им действовать самостоятельно, показали, что это не единственный тип нейронов, имеющих зрительно-моторные свойства1. К удивлению ученых, были обнаружены нейроны, активные и когда животное само выполняло моторный акт (например, брало пищу), и когда оно наблюдало за тем, как это делает экспериментатор. Такие нейроны были найдены в выпуклой части зоны F5 и названы зеркальными нейронами2.
Моторные свойства зеркальных нейронов аналогичны таковым у других нейронов зоны F5 в том, что они избирательно активируются в течение определенного моторного акта, но их зрительные свойства значительно отличаются.
В отличие от канонических нейронов, зеркальные не разряжаются при восприятии пищи или каких-либо иных трехмерных объектов, на их работу не влияет размер зрительного стимула. От чего их активность зависит — так это от наблюдения за определенными действиями, в которые вовлечена соответствующая часть тела (рука или рот), за взаимодействиями с объектами. Интересно, что копируемые моторные акты или нетранзитивные действия (не приложенные к определенному объекту), такие как поднятие или размахивание рукой, не вызывают ответа у таких нейронов. Также необходимо отметить, что активность зеркальных нейронов практически не зависит от положения в пространстве и расстояния от наблюдателя до наблюдаемого действия, хотя в некоторых случаях на нее влияет направление движения или используемая экспериментатором рука (правая или левая).
Если мы будем рассматривать зрительно кодируемый эффективный моторный акт в качестве критерия для классификации, зеркальные нейроны могут быть разделены на отдельные классы, сходные с теми, которые были выделены во второй главе при анализе моторных свойств у нейронов зоны F5. Итак, мы имеем «зеркальные нейроны хватания», «зеркальные нейроны удержания», «зеркальные нейроны манипулирования», а также «зеркальные нейроны размещения» (которые разряжаются, когда экспериментатор кладет объект на подставку) и «зеркальные нейроны взаимодействия с руками» (активные при наблюдении за рукой, движущейся в направлении другой руки, которая держит объект). Эта классификация показывает, что большинство зеркальных нейронов в F5 разряжаются при наблюдении за одним определенным типом действия (например, хватанием). Другие нейроны, похоже, не столь селективны, и разряжаются в ответ на наблюдение за несколькими — двумя или, реже, тремя — типами действий.
На рисунке 4.1 показано, как ведет себя типичный «зеркальный нейрон хватания». В условии А обезьяна наблюдает за тем, как экспериментатор поднимает с подноса кусочек пищи. Нейрон начинает разряжаться, как только рука экспериментатора достигает пищи и начинает принимать форму для ее захвата, и активность нейрона продолжается до конца действия. В условии Б обезьяна сама берет кусочек пищи, и в этом случае активность нейрона также коррелирует с изменением формы руки.
Сравнение разрядки нейрона при наблюдении и выполнении действия позволяет увидеть один из наиболее важных функциональных аспектов зеркальных нейронов: согласованность между кодированием выполнения моторного акта и наблюдением за выполнением его другим.
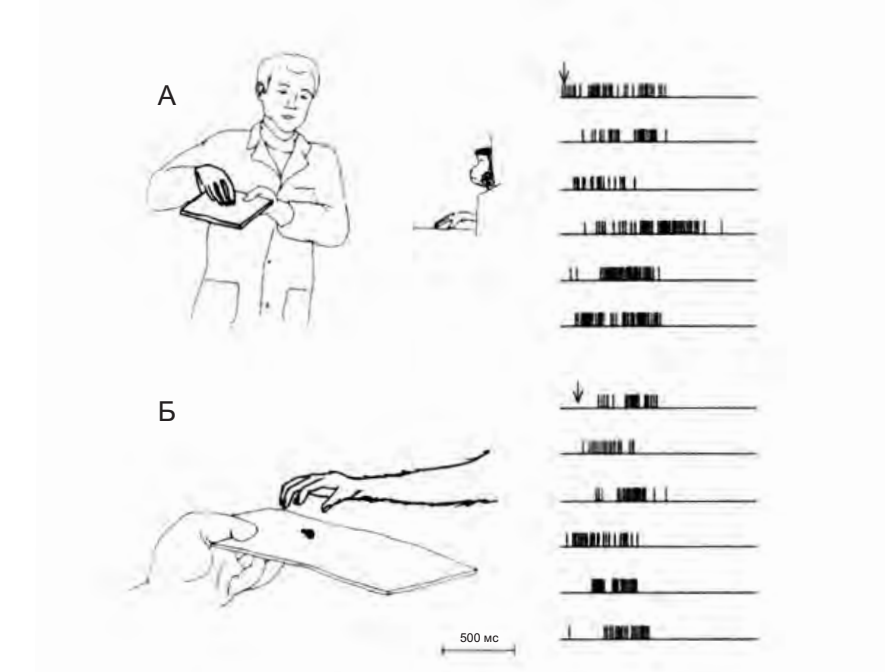
Рисунок 4.1. Зрительные и моторные ответы «зеркального нейрона хватания» (di Pellegrino et al. 1999)
Однако, у различных нейронов степень согласованности различается. Были выделены два основных типа: узко согласованные и широко согласованные. В нейронах с узкой согласованностью активность в ответ на выполняемое действие практически в точности повторяет активность при наблюдении. Пример приведен на рисунке 4.2. В условии А обезьяна наблюдает, как экспериментатор скручивает пальцами изюминку — вращая ее по часовой стрелке и против — как будто желая разорвать ее на две части. Нейрон разряжается только при вращении в одном из направлений. В условии Б экспериментатор и обезьяна держат одну и ту же изюминку, и нейрон разряжается, когда животное крутит ее в одном направлении, а экспериментатор — в другом, как бы пытаясь ее разорвать. В условии В обезьяна берет кусочек пищи с подноса при помощи движения захвата — нейрон не проявляет никакой активности.
В нейронах с широкой согласованностью кодируются наблюдаемое и выполняемое действие, которые явно связаны друг с другом, хотя могут не быть идентичны, и их связь может быть представлена на разном уровне обобщения. На самом деле, некоторые нейроны отвечают только на выполнение одного моторного акта (например, хватания) и на восприятие двух выполняемых актов (хватание и удержание).

Рисунок 4.2. Пример зеркального нейрона с узкой согласованностью. А — обезьяна наблюдает, как экспериментатор крутит изюминку в своих руках движениями, направленными в разные стороны; нейрон активен только при одном из двух направлений движения. Б — экспериментатор вращает кусочек пищи, который обезьяна держит в руке; животное вращает его в противоположном направлении. В — обезьяна берет пищу при помощи точного захвата. Для каждого условия показаны четыре серии записей активности нейрона. Стрелки показывают направление вращения в определенный момент записи (Rizzolatti et al. 1996a)
Другие кодируют только один выполняемый и наблюдаемый акт, но с разной степенью селективности. К примеру, рассмотрим нейрон, представленный на рисунке 4.3. Он разряжается, когда обезьяна наблюдает за экспериментатором, берущим объект при помощи точного захвата или захвата всей кистью, но отвечает на действия обезьяны, только когда она использует точный захват.
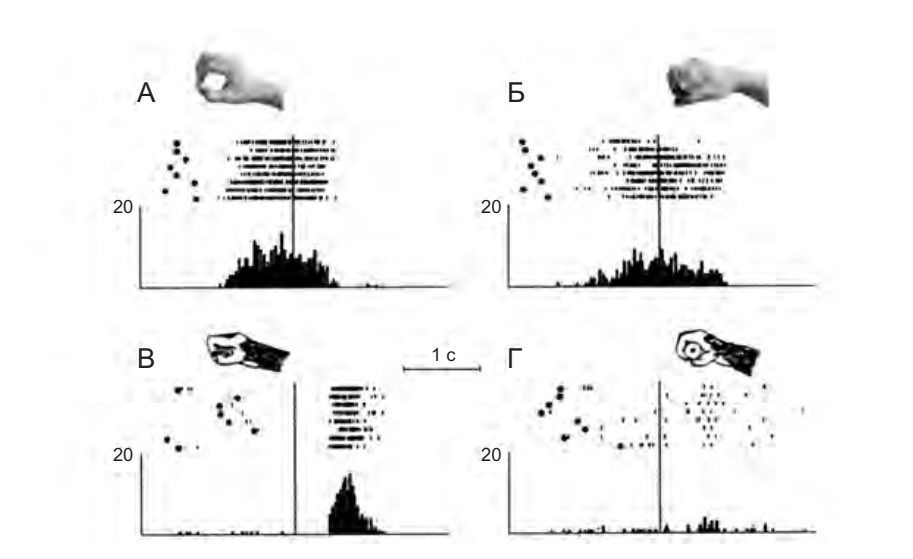
Рисунок 4.3. Пример широко согласованного нейрона. А — экспериментатор берет кусочек пищи, используя точный захват. Б — экспериментатор берет пищу всей кистью. В — обезьяна берет пищу точным захватом. Нейрон обладает высокой избирательностью к активному действию, но не к его наблюдению (Адаптировано из работы Gallese et al. 1996)
Кроме того, существуют нейроны, кодирующие действия зрительно — они разряжаются при выполнении другого действия, связанного с первым. Зеркальные нейроны такого типа разряжаются при наблюдении за тем, как экспериментатор кладет пищу на поднос, а также когда он ее берет. Функциональная роль и теоретическое значение таких нейронов подробно не обсуждались, однако, весьма вероятно, что их поведение есть последовательность организованных в цепь моторных актов. Мы обсудим этот вопрос в следующей части этой главы. С учетом различных типологий, нейроны с широкой степенью согласованности составляют примерно 70 % от общего числа зеркальных нейронов в коре обезьяны.

Коррадо Синигалья
Поедание и общение
До сих пор мы приводили примеры только тех зеркальных нейронов, которые связаны с движениями рук. Практически все первые исследования зеркальных нейронов были сконцентрированы вокруг дорзальной части зоны F5, большинство представленных в которой движений связаны с рукой. В первой главе мы отмечали, как при помощи микростимуляции нейронов электрическим током и регистрации их активности было показано, что движения рта также контролируются вентральной частью F5. Последние исследования показали, что нейроны этой области обладают зрительно-моторными свойствами так же, как и типичные зеркальные нейроны, и отвечают на выполнение движений ртом и наблюдение за движениями рта у других3.
На рисунке 4.4 показано выполнение экспериментатором и обезьяной некоторых действий, которые используются для оценки специфичности моторных и зрительных ответов нейронов животного. Рисунки вверху и в центре иллюстрируют два типичных транзитивных движения (то есть, действия, выполнение которых связано с объектом), которые включают заглатывание жидкости или твердой пищи. На рисунке внизу — нетранзитивное действие (вытягивание губ), которое наряду с чмоканием и оскаливанием зубов входит в репертуар коммуникативной мимики обезьян.
Большая часть зеркальных нейронов (около 85%) отвечают на восприятие действий, таких как хватание кусочка пищи губами, пережевывание или сосание его. Они были названы нейронами глотания. С функциональной точки зрения, эти нейроны сходны с зеркальными нейронами, кодирующими движения руки — они также разряжаются только при взаимодействии соответствующей части тела с объектом, а простое наблюдение покоящегося объекта или совершения нетранзитивного действия не вызывает никакого значимого ответа. Кроме того, большинство таких нейронов являются избирательными к определенному типу действия, и примерно треть из них — узко согласованные с ним (рисунок 4.5).
Зеркальные нейроны, отвечающие на восприятие коммуникативных актов, производимых при помощи рта, ведут себя иначе. На рисунке 4.6 приведены активности двух нейронов: в первом случае экспериментатор чмокает губами (А), вытягивает губы (Б), пьет из шприца (В). Только в условии А нейрон демонстрирует значимую активность. Но тот же самый нейрон становится активным, когда животное хватает пищу при помощи рта, при этом слегка вытягивая губы и язык (Г). Во втором случае экспериментатор вытягивает губы (А), держит пищу между зубами (Б), предлагает пищу обезьяне (В) — в этом случае нейрон также активируется только в условии А, и также разряжается, когда обезьяна выполняет типичное действие, связанное с глотанием — сосание сока из шприца (Г).
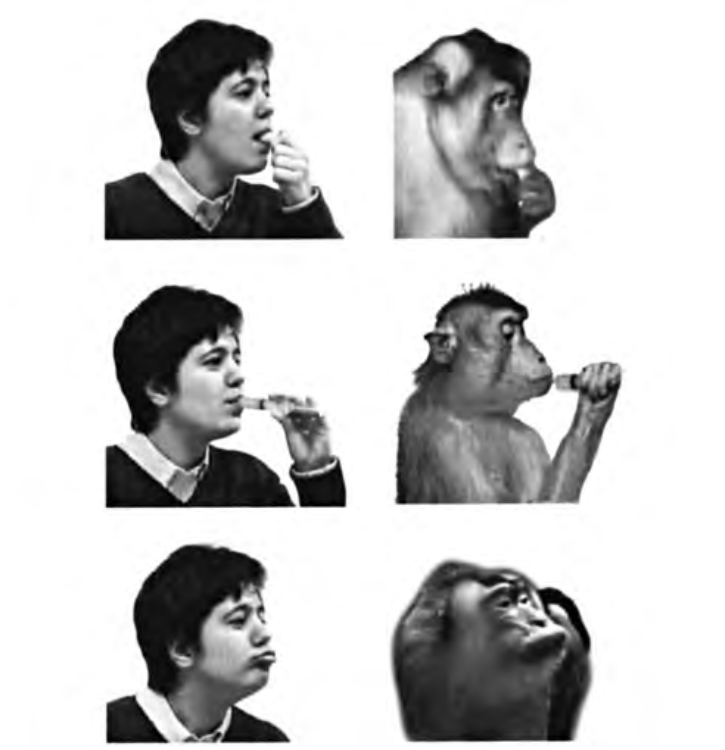
Рисунок 4.4. Примеры транзитивных и нетранзитивных действий экспериментатора и обезьяны, используемых для изучения зеркальных нейронов, кодирующих движения рта. Сверху вниз: захватывание пищи ртом; всасывание апельсинового сока из шприца; вытягивание губ.
Необходимо отметить, что в отличие от других зеркальных нейронов, коммуникативные нейроны отвечают при рассматривании нетранзитивных действий. Можно возразить, что эти ответы не связаны со зрительными стимулами как таковыми, но скорее с интерпретацией их животным как актов глотания. Другими словами, простое действие по вытягиванию языка экспериментатором вызовет моторную репрезентацию лизания у обезьяны. Такое объяснение кажется вполне разумным только потому, что оно придает законченный вид и единство теории о зеркальных нейронах. Однако, к сожалению, оно не объясняет того факта, что наблюдение за действиями глотания едва ли вызывает какую-либо активность у коммуникативных зеркальных нейронов.
Остается прояснить еще один важный вопрос: в отличие от зеркальных нейронов, которые связаны с движениями руки и поеданием пищи, коммуникативные зеркальные нейроны не демонстрируют согласованность между зрительным и моторным ответом.
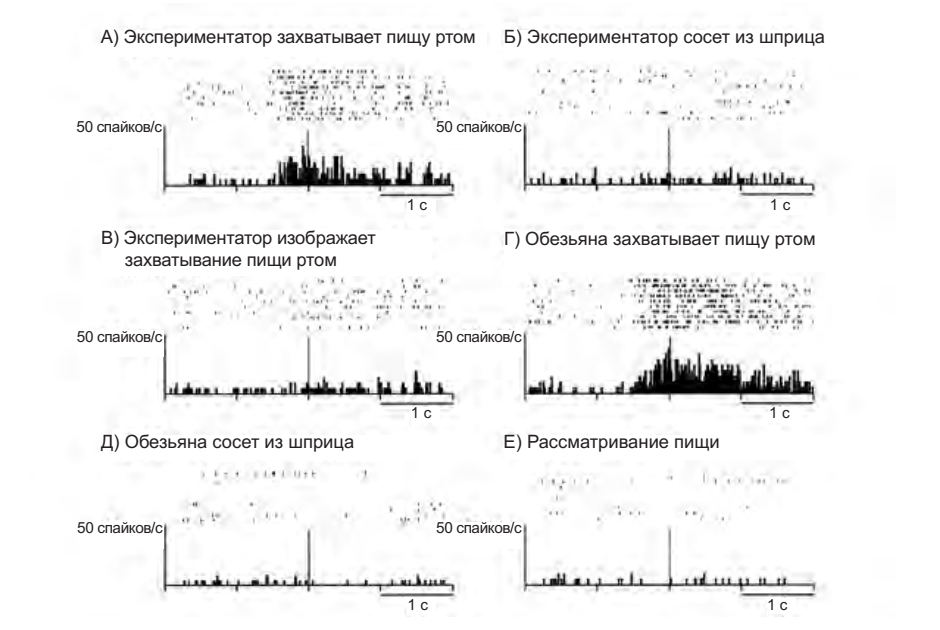
Рисунок 4.5. Пример активности «зеркального нейрона хватания ртом». А — экспериментатор наклоняется к подносу и берет с него пищу зубами. Б — экспериментатор подносит голову к шприцу, закрепленному на подставке, и сосет сок. В — экспериментатор изображает то же действие, что и в условии А, но без пищи. Г — экспериментатор подносит пищу к обезьяне, которая берет ее зубами и съедает. Д — экспериментатор подносит к обезьяне шприц с соком, обезьяна обхватывает губами шприц и сосет сок. Е — экспериментатор насаживает пищу на палочку и перемещает ее в поле зрения животного. На каждом графике представлены серии из десяти проб и усредненные гистограммы. Начало отсчета — момент, в который губы экспериментатора (зрительное кодирование) или обезьяны (моторное кодирование) касаются пищи, или в который пища попадает в поле зрения обезьяны. На рисунке В графики выровнены относительно момента окончания движения. По абсциссе — время в секундах, по ординате — количество спайков за каждые 20 мс
Только первый из них по своей природе коммуникативный, тогда как второй связан с глотанием. Если мы рассмотрим чисто моторные свойства, то в большинстве случаев найдем высокую корреляцию между наблюдаемым и выполняемым действиями: нейрон, который разряжается при наблюдении за вытягиванием губ, активен и при действиях обезьяны, требующих сходных движений (например, сосании сока из шприца), но не отвечает при выполнении других действий. То же самое верно и для чмокания губами. Однако, необходимо помнить, что значения этих действий различаются. Убедить обезьяну выполнять чисто коммуникативные действия в экспериментах с регистрацией активности отдельных нейронов — задача не из легких.
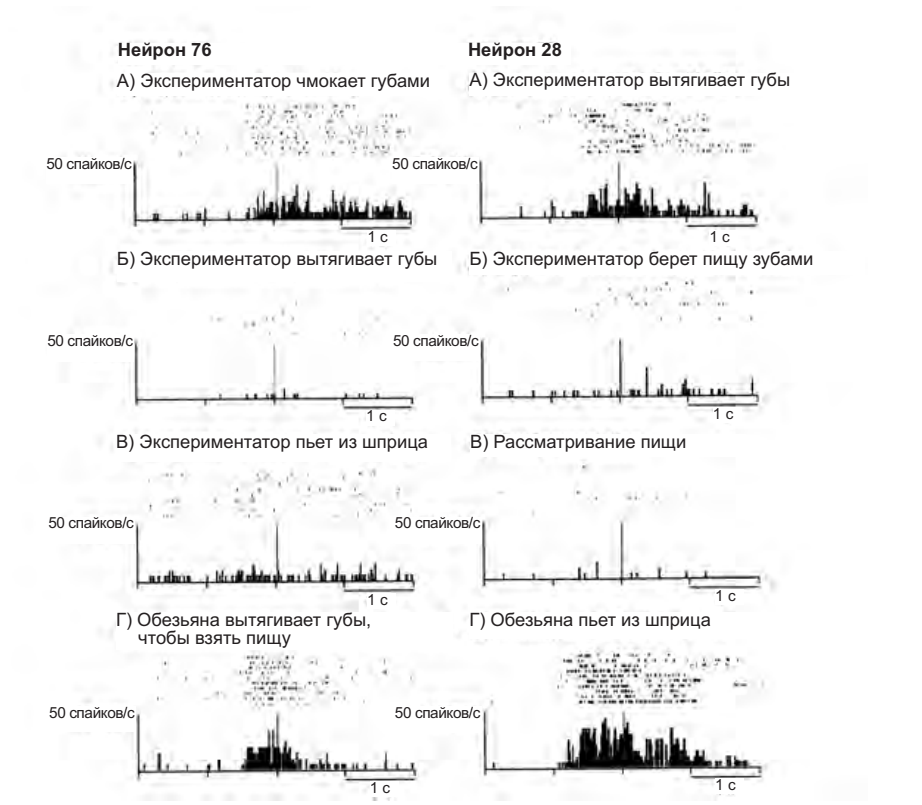
Рисунок 4.6. Примеры двух коммуникативных зеркальных нейронов. Активность в отдельных пробах и результирующие гистограммы выровнены относительно пика действия. Нейрон 76: А — экспериментатор чмокает губами перед обезьяной, Б — экспериментатор вытягивает губы, В — экспериментатор придвигается к закрепленному шприцу с соком и втягивает содержимое, Г — экспериментатор предлагает пищу обезьяне, которая вытягивает губы и хватает ее. Нейрон 28: А — экспериментатор вытягивает губы, глядя на обезьяну, Б — экспериментатор придвигается к пище, закрепленной на подставке, хватает ее зубами и держит, В — экспериментатор помещает палочку с кусочком пищи перед обезьяной, Г — экспериментатор предлагает обезьяне попить из шприца, обезьяна пьет (Ferrari et al. 2003)
Тем не менее, в редких случаях, когда это удавалось, были зарегистрированы четкие нейронные ответы на коммуникативные действия (рисунок 4.7). Эти результаты вселяют в нас надежду, что такая активность свойственна многим, а не только нескольким, нейронам.
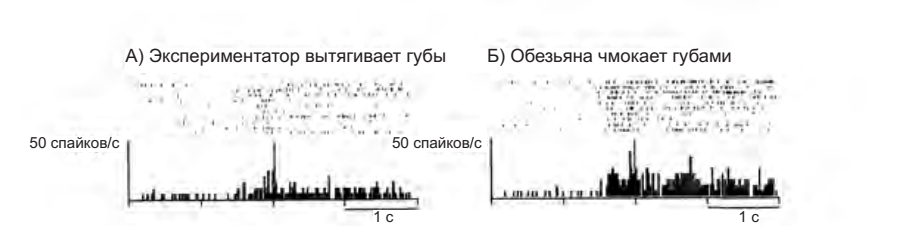
Рисунок 4.7. Пример коммуникативной зеркальной системы, кодирующей вытягивание губ. А — экспериментатор вытягивает губы, глядя на обезьяну. Б — обезьяна отвечает на чмокание экспериментатора чмоканием (Ferrari et al. 2003)
Если оставить в стороне трудности, с которыми мы можем столкнуться в экспериментальных условиях, тот факт, что действия поедания пищи и общения имеют общий нейронный субстрат, кажется нам исключительно интересным, особенно в свете некоторых этологических исследований на приматах4. Коммуникативные действия, такие как чмокание или вытягивание губ, сформировались на основе репертуара движений, изначально связанных с поеданием и грумингом. Хорошо известно, что у приматов груминг является одним из важнейших способов выражения дружелюбного отношения и солидарности с сообществом: он способствует образованию группы, а когда она становится слишком большой — разделению ее на коалиции с целью защиты слабейших членов от агрессии других животных. Когда обезьяна начинает чистить и вычесывать насекомых из шкуры другой обезьяны, она часто сопровождает или предваряет свои первые движения чмоканием. Кормление также предваряется чмоканием, но с другим звуком, словно обезьяны хотят подчеркнуть различия между этими действиями. Таким образом, чмокание в отсутствие груминга оказывается формой ритуализированного моторного акта, который преобразует функции, связанные с объектом, в коммуникативные. То же самое верно и для таких движений, как вытягивание губ или языка. В таком контексте открытие коммуникативных зеркальных нейронов в такой зоне, как F5, вместе с несомненным отсутствием согласования между зрительным и моторным ответами, отражает первичный процесс обработки в коре коммуникативных сигналов, которые еще не полностью отделились от пищевого поведения (то есть, от транзитивных действий по поднесению пищи ко рту и ее заглатыванию).
Связи верхней височной извилины и нижней теменной коры
Как мы видели во второй главе, большая часть зрительной информации, поступающей к каноническим нейронам F5, приходит к ним из передней межтеменной зоны (AIP) [The anterior intraparietal]. Теперь перед нам стоит вопрос о том, какие участки коры поставляют информацию в зеркальные нейроны.
Более пятнадцати лет назад Дэвид И. Перретт и его коллеги5 показали в эксперименте на обезьянах, что передняя часть верхней височной борозды (STS) содержит нейроны, селективно отвечающие на восприятие большого ряда движений тела, выполняемых другим индивидом — некоторые нейроны были активны, когда обезьяна наблюдала за движениями глаз или головы, другие отвечали на движения туловища и ног (хождение), а третьи снова кодировали определенные взаимодействия рук с объектом.
Зрительные свойства нейронов последнего типа кажутся весьма сходными с таковыми у зеркальных нейронов зоны F5. И те, и другие кодируют в значительной мере сходные типы наблюдаемых движений на разном уровне обобщения (например, хватание рукой, использование точного захвата и т. д.) и неактивны при наблюдении за тем, как экспериментатор выполняет нетранзитивные действия или изображает транзитивные действия в отсутствие объектов.
Однако, можно отметить одно существенное расхождение между ними: в отличие от зеркальных нейронов зоны F5, нейроны STS являются чисто зрительными и не принимают участия в кодировании движений. Следовательно, они не обладают свойством совмещения зрительных и моторных функций, что является важной характеристикой зеркальных нейронов.
Нейроны верхней височной борозды чрезвычайно интересны не только из-за их свойств, но также потому, что они помогают нам понять, каким образом возможно возникновение таких сложных по своим функциям клеток как зеркальные нейроны. Они демонстрируют, как происходит кодирование биологических движений, выполняемых другими индивидами, в специфической корковой системе, и как процесс идентификации этих движений начинается в зрительной коре. Следующий шаг кажется довольно простым — предположить, что зрительная информация передается в моторную кору, обеспечивая зрительные свойства моторных нейронов.
Далее встает вопрос о том, как информация передается из STS в F5. С анатомической точки зрения, STS не имеет прямых проекций в вентральную премоторную кору. Однако, она имеет мощные связи с нижней теменной корой и участками префронтальной коры6. Зрительная информация о наблюдаемых действиях может, таким образом, достичь зоны F5 по одному из этих путей. Скорее всего, второй из них не играет важной роли — известно, что связи F5 с префронтальной корой, получающей информацию от STS, не очень сильны. С другой стороны, F5 связана с ростральной частью нижней теменной коры, в которую входят зоны PF и PFG5.
Более того, функции нейронов области PF—PFG говорят о том, что этот комплекс может рассматриваться как мост между зонами STS и F5. В работах Яри Хюваринена и его коллег6, проведенных в 80-е годы, было показано, что нейроны этого комплекса отвечают на сенсорные (соматосенсорные и зрительные) стимулы, и что около трети из них также активируются при выполнении произвольных движений руки и рта. Позднейшие исследования7 показали, что примерно 40 % нейронов, отвечающих на зрительные стимулы, активны во время наблюдения за моторным актом, в который включена рука — например, хватанием, удержанием, дотягиванием, — и что еще более важно, большая часть нейронов (около 70 %) обладает моторными свойствами, отвечая при выполнении обезьяной действий при помощи рук, рта или того и другого. Это теменные зеркальные нейроны.
Как и нейроны зоны F5, теменные зеркальные нейроны не отвечают на простое наблюдение за пассивным субъектом или покоящимся объектом действия, и даже на изображение действия в отсутствие объекта. Половина из них избирательно кодирует только один тип моторного акта, другая половина избирательна к двум видам действий (например, хватание и отпускание). Учитывая связи между наблюдаемым и выполняемым действиями, они работают так же, как и зеркальные нейроны в F5: некоторые из них являются узко согласованными, большинство — широко согласованными.
Функции зеркальных нейронов
Теперь нам предстоит прояснить функциональное значение зеркальных нейронов, зарегистрированных в зонах F5 и PF—PFG. На первый взгляд, мы можем приписать активность этих нейронов при восприятии обезьяной действий, выполняемых другими (в нашем случае, экспериментатором), неспецифическим факторам, таким, как внимание или ожидание пищи, либо подготовке к действию, которая обеспечивает повторение животным увиденных действий настолько быстро, насколько это возможно, чтобы быть готовым к возможному соперничеству с окружающими. Если бы дело обстояло так, то зеркальные нейроны либо были бы лишены всякой специфичности, либо просто представляли бы собой определенную категорию «подготовительных нейронов», которые широко распространены в премоторной коре и которые активируются перед эффективным выполнением действия.
Более внимательный взгляд покажет, что ни одну из этих гипотез нельзя принять. Избирательность ответа и зрительно-моторное согласование, обнаруженные у большинства зеркальных нейронов, не могут быть отнесены на счет поведения, связанного с ожиданием пищи или какой-либо другой формы поощрения. Эксперимент, представленный на рисунке 4.8, подтверждает это. В условиях А и Б обезьяна, у которой регистрировалась активность нейронов, наблюдала за другой обезьяной или экспериментатором, когда они брали пищу при помощи рук. В условии В обезьяна сама выполняла такое действие. Зеркальные нейроны разряжались в условиях А и Б, несмотря на то, что в обоих условиях обезьяна не могла дотянуться до пищи или получить какую-либо другую награду.
Объяснение активности зеркальных нейронов как формы подготовки к действию не является удовлетворительным. На рисунке 4.8 можно увидеть, что в то время как первая обезьяна наблюдает за второй, хватающей пищу, ей нет никакого смысла готовиться к действию — она не может дотянуться до пищи. Кроме того, читатель вспомнит, что ни в одном из вышеописанных экспериментов разрядка зеркальных нейронов при наблюдении за выполнением действия не сопровождалась последующим самостоятельным его выполнением. Мы не должны забывать, что зеркальные нейроны никогда не активировались, если пища предлагалась обезьяне на таком расстоянии, что до нее можно было дотянуться. Если бы ответ нейронов был связан с подготовкой к действию, они бы разряжались перед тем, как обезьяна сама его выполняла.
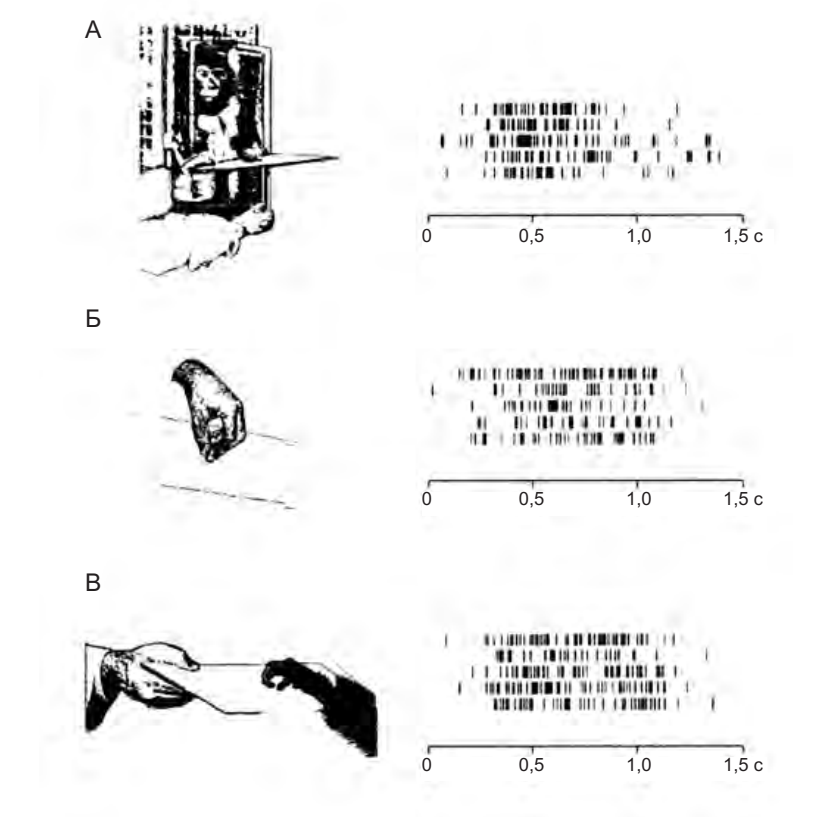
Рисунок 4.8. Активация «зеркального нейрона хватания». А — обезьяна, активность нейронов которой регистрируется, наблюдает за другой обезьяной, берущей пищу при помощи рук. Б — обезьяна смотрит, как экспериментатор берет пищу. В — обезьяна сама выполняет это действие. Для каждого условия показаны по пять записей активности нейрона. Спонтанная активность практически отсутствовала (Rizzolatti et al. 1996a)
Несколько лет назад Марк Жанро в своей статье, посвященной воображаемым действиям, предложил другую (более сложную) интерпретацию функций зеркальных нейронов8. Представьте себе урок музыки и ученика, внимательно следящего за тем, как его преподаватель исполняет сложный пассаж на скрипке, который ему нужно будет повторить после того, как учитель закончит играть. Для этого ученику нужно построить моторный образ быстрых движений пальцев маэстро. Далее, согласно Жанро, те же нейроны, которые отвечают за построение моторного образа, будут активны и во время подготовки с ученика и исполнения пьесы. Другими словами, активация зеркальных нейронов позволяет создавать «внутренние моторные репрезентации» наблюдаемых действий, на которых основана способность к обучению при помощи подражания.
Предположение Жанро чрезвычайно важно и согласуется с результатами экспериментов, которые мы рассматривали ранее. Тесная связь между зрительными и моторными ответами зеркальных нейронов свидетельствует о том, что в то время как индивид наблюдает за выполнением действий другими, в его мозге выполняется потенциальный моторный акт, по всем своим характеристикам сходный с тем, который спонтанно активируется при подготовке и выполнении реального действия. Различие лишь в том, что в одном случае это действие остается на стадии потенциального (в виде «внутренней моторной репрезентации»), а в другом оно переводится в последовательность конкретных движений. Однако, в одном вопросе мы не можем согласиться с Жанро, а именно, мы не верим в то, что основной функцией зеркальных нейронов является обеспечение имитации действий.
Далее мы более глубоко проанализируем широкий ряд феноменов, которые часто рассматриваются как — и часто путаются с — имитацией; также мы оценим, до какой степени способность человека обучаться выполнению действий после наблюдения за тем, как их выполняют другие, зависит от функционирования системы зеркальных нейронов. В любом случае, за последние годы этологи все больше стали утверждаться во мнении, что имитация в правильном смысле этого слова — прерогатива человека и (возможно) человекообразных обезьян, но не макак, на которых проводились описанные нами эксперименты9. Поэтому мы не можем полностью согласиться с интерпретацией Жанро: функции зеркальных нейронов зоны F5 и комплекса PF—PFG имеют более древнее эволюционное происхождение, и на основании приведенных примеров можно сказать, что изначально они были включены в понимание значений «моторных событий» — то есть, действий, производимых другими10.
Используемый нами термин «понимание» не обязательно означает, что наблюдатель (в данном случае, обезьяна) имеет эксплицитное знание или даже осознает то, что наблюдаемое и выполняемое действия идентичны или очень похожи. То, что мы утверждаем, намного проще — мы говорим о способности незамедлительно распознавать определенные типы действий в наблюдаемом «моторном событии», типы действий, которые характеризуются определенным способом взаимодействия с объектами. Это нужно для того, чтобы отличить это действие от других и затем использовать это знание для выбора наиболее подходящего ответа. Таким образом, то, что ранее было сказано относительно канонических нейронов зоны F5 и зрительно-моторных нейронов передней межтеменной коры (AIP), остается верным и в данном случае: зрительный стимул начинает обрабатываться незамедлительно, начиная с соответствующего моторного акта, даже если последний не был реально выполнен. Есть только одно важное различие — в случае с зеркальными нейронами зрительный стимул это не объект или его движения, но движения другого индивида, направленные на хватание, удержание, манипулирование объектом.
Как и в случае с объектами, эти движения приобретают значение для индивида благодаря словарю моторных актов, который регулирует его способность выполнять действия. У обезьяны этот словарь включает захватывание пищи, ее удержание, поднесение ко рту, и так далее.
Вот почему когда она видит, как экспериментатор сгибает свою руку для точного захвата и протягивает ее по направлению к пище, она незамедлительно воспринимает значение этих «моторных событий» и интерпретирует их в терминах целенаправленного действия.
Зрительная репрезентация и моторное понимание действия
Однако, на это имеется и очевидное возражение: как уже обсуждалось ранее, нейроны, избирательно отвечающие на наблюдение за движениями тела других, и в некоторых случаях — за взаимодействием руки и объекта — были обнаружены в передней части верхней височной борозды (STS). Мы отмечали, что эта область связана с зрительной — височной и затылочной — корой и формирует систему, во многом параллельную вентральному потоку (см. рис. 2.7). Какой же смысл тогда говорить о системе зеркальных нейронов, которая обеспечивает формирование в мозге наблюдателя образа действия, производимого другим, и описание его в терминах его собственного моторного акта? Не будет ли намного проще предположить, что понимание действий других основано на чисто зрительных механизмах анализа и синтеза различных элементов, составляющих наблюдаемое действие, без какого-либо моторного вмешательства со стороны наблюдателя?
Перретт и его коллеги показали11, что зрительное кодирование действий в передних областях STS производится на чрезвычайно высоком уровне сложности. Для примера, в этой зоне имеются нейроны, способные объединять получаемую при наблюдении информацию о направлении взгляда и выполняемых движениях. Такие нейроны активируются, только когда обезьяна наблюдает, как экспериментатор берет предмет, на который он при этом смотрит. Если экспериментатор переведет свой взгляд в другую сторону, наблюдение за его дейcтвиями не вызовет никакой значимой активации нейрона. Однако, мы должны выяснить, можно ли эту избирательность — или, в более общем смысле, способность связывать различные воспринимаемые аспекты выполняемого действия — обозначать термином «понимание». Особенности моторной активации нейронов зон F5 и PF—PFG привносят то, что не может быть выведено из чисто зрительных свойств нейронов STS, и без чего взаимосвязь воспринимаемых аспектов действия будет в лучшем случае случайной, ничего не значащей для наблюдателя.
С точки зрения моторных свойств, связь между действием по дотягиванию до предмета и направлением взгляда, в действительности, с не является случайной: еще в колыбели мы научаемся тому, что лучший способ получить объект — уставиться на него. Как и все успешные стратегии, эта становится частью нашего словаря действий, и, таким образом, когда мы видим, как кто-либо воспроизводит это действие, наша моторная система входит в резонанс с ним, благодаря чему мы распознаем интенциональный аспект движений другого и понимаем тип его действий.
Зрительно-моторные свойства зеркальных нейронов позволяют им координировать зрительную информацию с моторным знанием. Активация зеркальных нейронов как моторных во время выполнения действия характеризуется не только тем, что кодируются тип, модальность и временная развертка действия, но также обеспечивается контроль за его выполнением. А контроль за двигательным процессом невозможен без механизма антиципации, и, таким образом, каждый контролируемый процесс определяет корреляцию между соответствующей нейронной активностью и эффектами, которые она порождает. В конкретном случае зон F5 и PF—PFG взаимная координация позволяет сформировать базовое моторное знание о значениях действий, кодируемых различными нейронами — знание, которое может быть использовано как во время выполнения действия, так и при наблюдении за его выполнением другими.
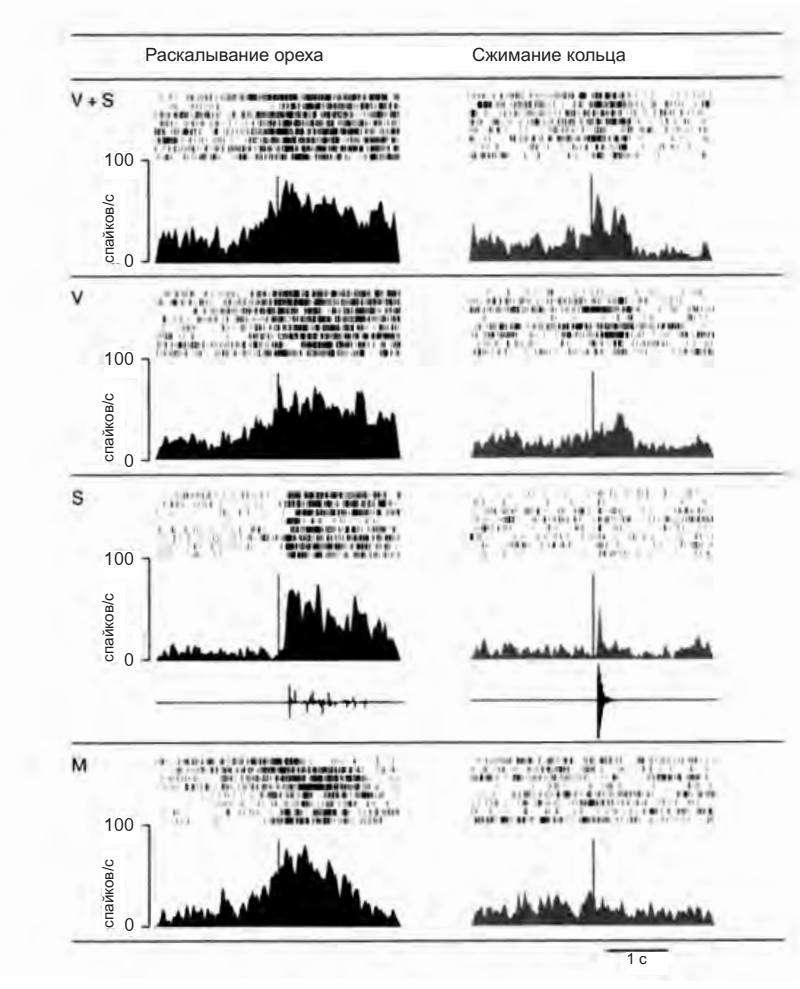
Рисунок 4.10. Два «аудиовизуальных» зеркальных нейрона F5. Вертикальными линиями на гистограммах отмечено начало звука в условиях «зрение и слух» (V + S) и «слух» (S). В условии «зрение» (V) они обозначают тот момент, в который мог бы начаться звук, если бы он предъявлялся. В чисто моторном условии (M) они отмечают момент, когда обезьяна дотрагивается до объекта (Kohler et al. 2003)
Активация той же самой совокупности нейронов говорит о том, что понимание действий других предполагает наличие у наблюдателя того же самого знания о принципах выполнения моторных актов, которое регулирует его собственное выполнение действий.
В некоторых недавних исследованиях была подтверждена гипотеза о том, что моторное знание играет фундаментальную роль в обработке сенсорной информации — до той степени, что без нее вообще было бы сложно говорить о понимании действия12.
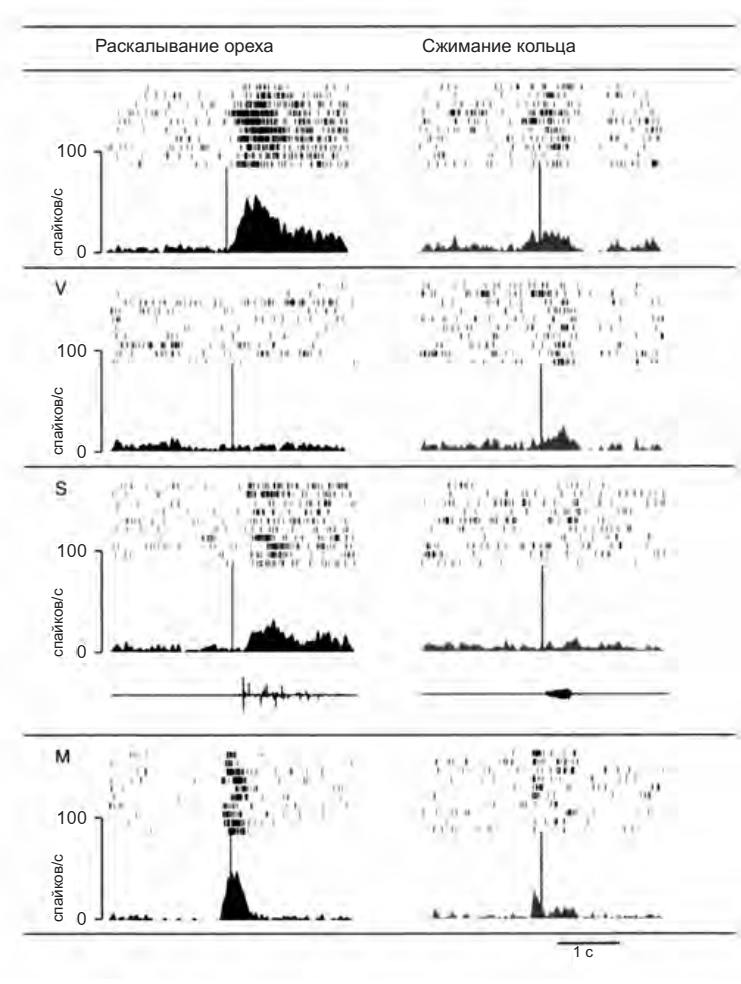
Рисунок 4.10. (продолжение)
Мария Алессандра Умильта и ее коллеги показали, что большинство нейронов зоны F5 отвечают при выполнении действий экспериментатором, независимо от того, что их финальная фаза (то есть, наиболее важный этап — непосредственное взаимодействие руки с объектом) скрыта от обезьяны. Эксперимент представлен на рисунке 4.9 (см. цв. вклейку). Активность отдельных нейронов регистрировалась в четырех различных условиях: А — обезьяна наблюдает за тем, что делает экспериментатор, и полностью видит последнюю часть действия (захват объекта); Б — она видит только начало действия, так как его окончание скрыто экраном; В и Г — условия, аналогичные условиям А и Б, за исключением того, что экспериментатор изображает действие при отсутствии объекта.
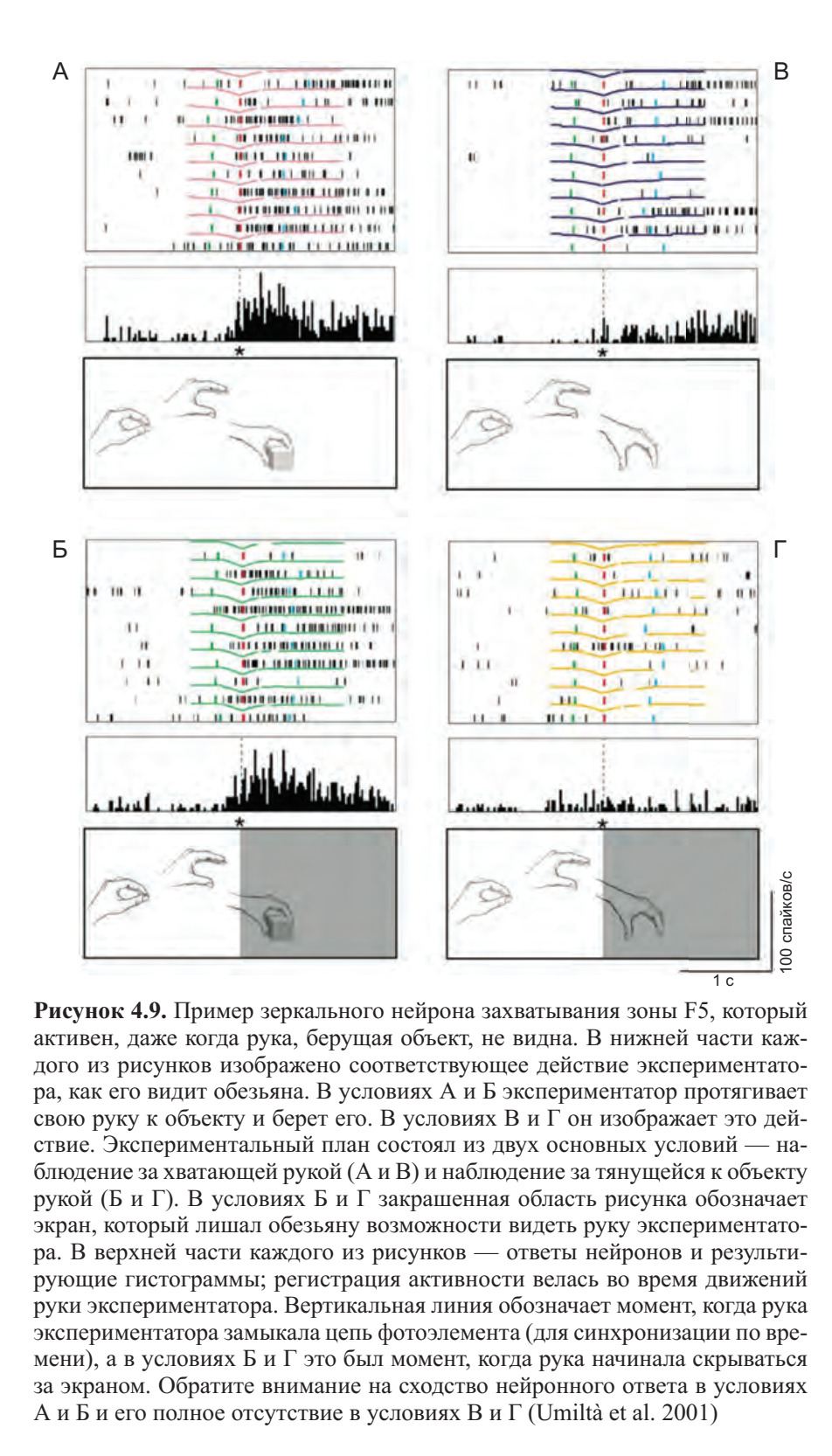 Данные регистрации активности нейронов показали: тот факт, что обезьяна не могла видеть финальную часть действия, никоим образом не изменял паттерн активации нейрона по сравнению с условием, в котором рука экспериментатора была видна на протяжение всего действия. В условии Б обезьяна видела, что объект закрывается экраном. Однако, нельзя говорить, что ответ нейрона есть всего лишь «реминисценция объекта», так как в таком случае разрядка нейрона началась бы, как только объект попадал в поле зрение, а это было не так. Напротив, поведение нейрона говорит о том, что тот же самый моторный акт, только потенциальный, происходит и когда обезьяна наблюдает действие полностью, и когда видит только его начало. В действительности, именно этот потенциальный моторный акт («внутренняя моторная репрезентация») позволяет обезьяне восстановить отсутствующую часть действия и распознать его значение по увиденной части.
Данные регистрации активности нейронов показали: тот факт, что обезьяна не могла видеть финальную часть действия, никоим образом не изменял паттерн активации нейрона по сравнению с условием, в котором рука экспериментатора была видна на протяжение всего действия. В условии Б обезьяна видела, что объект закрывается экраном. Однако, нельзя говорить, что ответ нейрона есть всего лишь «реминисценция объекта», так как в таком случае разрядка нейрона началась бы, как только объект попадал в поле зрение, а это было не так. Напротив, поведение нейрона говорит о том, что тот же самый моторный акт, только потенциальный, происходит и когда обезьяна наблюдает действие полностью, и когда видит только его начало. В действительности, именно этот потенциальный моторный акт («внутренняя моторная репрезентация») позволяет обезьяне восстановить отсутствующую часть действия и распознать его значение по увиденной части.
Исследование, проведенное Эвелин Келер и ее коллегами13, является еще одним доказательством в пользу гипотезы, что активация моторных нейронов отражает значение наблюдаемого действия и не зависит только от его зрительно воспринимаемых характеристик. Авторы выделили особый тип бимодальный зеркальных нейронов в зоне F5 (аудиовизуальные нейроны), которые активируются и при наблюдении обезьяной за действиями экспериментатора, сопровождающимися характерными звуками, и когда животное слышит эти звуки без зрительного восприятия действия.
На рисунке 4.10 представлена активность двух таких нейронов: они не только являются селективными к определенному действию (например, раскалыванию земляного ореха), но также их ответы в различных экспериментальных условиях (зрительное и слуховое восприятие; только зрение; только слух; самостоятельное выполнение действия) совершенно совпадают. Это означает, что вызываемый потенциальный моторный акт всегда один и тот же, тогда как сенсорная информация может меняться в зависимости от конкретной ситуации.
Зрительные характеристики действия оказываются релевантными только до той степени, пока они облегчают понимание действия. Однако, если для этого достаточно других факторов (например, характерного звука), зеркальные нейроны способны кодировать действия экспериментатора даже в отсутствие зрительных сигналов.
Мелодия действия и понимание намерений
Демонстрация того, что зеркальные нейроны зон F5 и PF—PFG обеспечивают понимание действий других, не должна пониматься как единственная функция зеркального механизма. Далее мы приведем целый ряд иллюстраций разнообразных функций зеркальных нейронов и карту корковых зон, включенных в эту систему как у человека, так и у других приматов. Однако, это ни в коей мере не отменяет значимости механизма понимания действий, обеспечиваемого работой нейронов зон F5 и PF—PFG и отмеченного согласованностью их сенсорных и моторных ответов. Это, как мы уже видели, является имплицитной формой понимания их происхождения и значения — оно не связано с конкретной сенсорной модальностью восприятия действия, но ограничено имеющимся словарем моторных актов, который выполняет функцию регуляции и контроля моторного компонента действия.
Следовательно, полная или частичная замена зрительной информации на слуховую не изменяет ход событий. Даже если она будет в высшей степени специфичной и достигнет высокой стадии обработки, как происходит в нейронах верхней височной извилины, все равно в ней будет отсутствовать то моторное значение, которое может быть сформировано только благодаря двусторонним связям между нейронами F5 и системы PF—PFG, где воспринятые при помощи зрения или слуха действия других приобретают специфическое значение или становятся целенаправленными, или интенциональными, моторными актами.
Таким образом, моторное знание о наших собственных действиях есть необходимое и достаточное условие незамедлительного понимания действий других. Как будет показано далее, это знание фундаментально значимо для построения базового понимания целенаправленности, потому что оно обеспечивает не только отдельные моторные акты, которые мы рассматривали до сих пор, такие как хватание, удержание, разрывание, но также и соединение их в более сложные действия.
Этот аспект моторной организации был недавно изучен Леонардо Фогасси и его коллегами14, которые регистрировали активность ряда зеркальных нейронов в теменной коре (рис. 4.11 А) при хватании обезьяной объекта. В эксперименте было два условия: в первом обезьяна начинала движение рукой из заранее определенной точки до кусочка пищи, расположенного перед ней, чтобы его схватить и донести до рта. Во втором условии исходное положении руки было таким же, но вместо того, чтобы положить пищу себе в рот, обезьяна клала его в контейнер (рис. 4.11 Б).
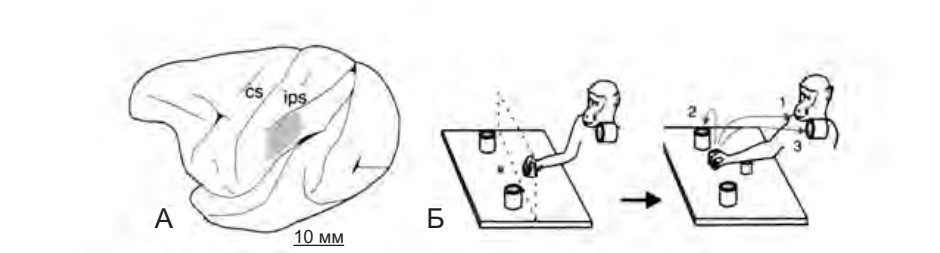
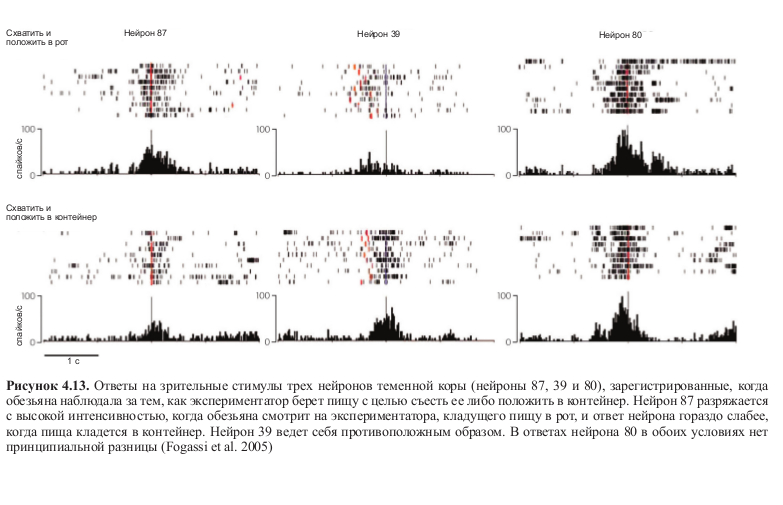
Рисунок 4.11. А — латеральный срез мозга обезьяны. Закрашенный участок — часть нижней теменной коры, активность нейронов которой регистрировалась в эксперименте. Б — условия эксперимента. Контейнер располагался либо рядом с пищей, либо на плече у обезьяны. Так как скорость движения по дотягиванию и захвату пищи определялась последующим моторным актом, при помощи изменении позиции контейнера (и, следовательно, траектории движения) можно было определять, зависит ли избирательность нейронов от особенностей выполнения движения или от конечной цели действия. Результаты эксперимента показали, что в действительности селективность нейронов определяется целью действия (Fogassi et al. 2005).
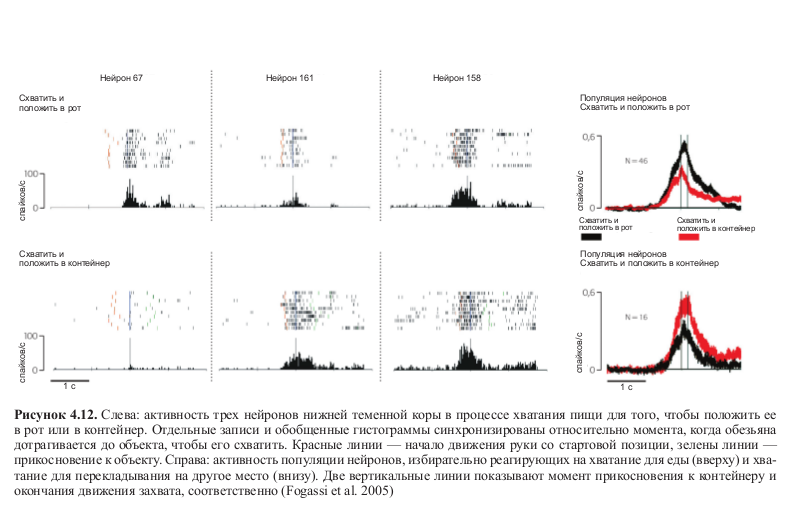
Результаты этих экспериментов показали, что большинство нейронов, активность которых регистрировалась, разряжались по-разному в зависимости от того, какое действие следовало за хватанием пищи — помещение ее в рот или в контейнер (см. табл. 4.1). На рисунке 4.12 (см. цв. вклейку) показана активность трех нейронов: можно увидеть, что два из них разряжаются по-разному в зависимости от того, какое действие следует за хватанием пищи. Третий нейрон кодирует движение хватания независимо от цели действия.
 Была также проведена серия контрольных экспериментов для того, чтобы выяснить, чем еще может быть объяснена избирательность нейронов к определенному действию («положить в рот» и «переместить с одного места на другое») — типом объектов (пища и несъедобные предметы) или силой, с которой обезьяна хватает объект.
Была также проведена серия контрольных экспериментов для того, чтобы выяснить, чем еще может быть объяснена избирательность нейронов к определенному действию («положить в рот» и «переместить с одного места на другое») — типом объектов (пища и несъедобные предметы) или силой, с которой обезьяна хватает объект.
Эти альтернативные интерпретации были отвергнуты по результатам контрольных экспериментов. Кроме того, изучение кинематического рисунка (скорость, ускорение, и т. д.) движений по дотягиванию и хватанию объекта показало, что моторные характеристики выполнения действия не влияют на избирательность нейронов к тем или иным действиям.

Таблица 4.1. Нейроны нижней теменной коры, активность которых изучалась при выполнении действия по захвату пищи для того, чтобы положить ее в рот или в контейнер
Читателю может показаться противоречащим здравому смыслу предположение о том, что моторные нейроны теменной коры настроены на специфические действия. Не слишком ли это расточительно — иметь «нейроны хватания» для каждого конкретного действия? Не было бы более экономным иметь возможность подключать к работе любые из них, когда нам нужно что-нибудь схватить? Ответ на этот вопрос следует искать в фундаментальных свойствах моторной организации: типичным для действий человека и многих животных является плавность действий. «Нейроны хватания» встраиваются в заранее сформированные нейронные цепочки, обеспечивающие выполнение всего действия, таким образом, что каждый нейрон кодирует хватание, но в то же время, будучи включенным в последовательно выполняемый моторный акт, обеспечивает плавность движений.
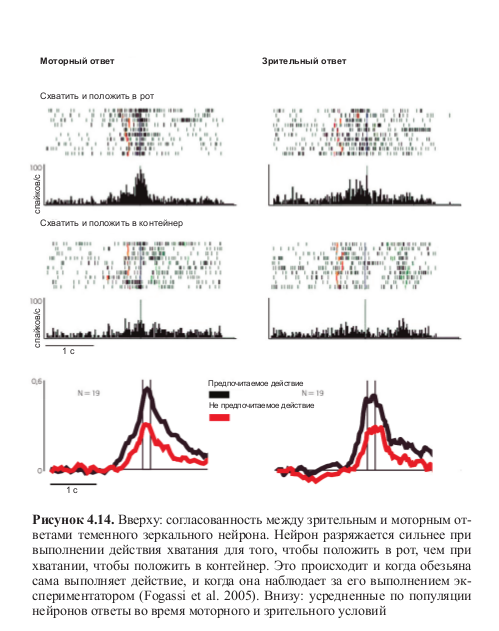
Но еще более интересным может показаться другой аспект исследования Фогасси и коллег: некоторая форма избирательности, сходная с избирательностью моторной, проявлялась в тот момент, когда обезьяна просто наблюдала за тем, как экспериментатор выполняет ту же последовательность действий. В этом случае нейроны также разряжались по-разному в зависимости от типа действия, в который было встроено кодируемое ими движение. Более того, эти избирательные нейроны также демонстрировали высокую согласованность моторного и зрительного ответов (табл. 4.2, рис. 4.13 и 4.14, см. цв. вклейку).

Таблица 4.2. Нейроны нижней теменной коры, активность которых изучалась при наблюдении за действием по захвату пищи для того, чтобы положить ее в рот или в контейнер
Стоит отметить, что в обоих случаях — когда обезьяна сама выполняет действие и когда она смотрит на выполнение его экспериментатором — нейроны начинают разряжаться, как только рука (обезьяны или экспериментатора) принимает форму, необходимую для захвата пищи или объекта. На самом деле, не так уж и удивительно то, что нейроны начинают кодировать значение целенаправленного действия с самого первого движения. Когда обезьяна протягивает руку по направлению к пище, она уже знает о том, что она собирается делать — донести эту пищу до рта или положить ее в другое место. Даже если намерение животного становится явным только после полного развертывания моторного поведения, оно все равно определяет начальный этап действия. Кроме того, гипотеза об организации моторных актов в специфические моторные цепочки также подтверждается организацией соматосенсорных рецептивных полей нейронов теменной коры. То, что многие теменные нейроны, отвечающие на пассивное сгибание руки, имеют еще и тактильные рецептивные поля, расположенные вокруг рта, свидетельствует о следующем: эти нейроны будут обеспечивать раскрытие рта, если животное будет хватать и поднимать объект15.
 Но эта ситуация меняется, когда обезьяна наблюдает за тем, как экспериментатор берет рукой пищу или другой объект. Однако, тот факт, что зрительный стимул активирует тот же самый нейронный паттерн — то есть, тот же набор потенциальных моторных актов, который отвечает за работу всей моторной цепочки — говорит о том, что обезьяна с самого начала способна ухватить реальную динамику целенаправленных движений, за которыми она наблюдает, и предугадать, чем завершится действие экспериментатора, как только он начнет движение.
Но эта ситуация меняется, когда обезьяна наблюдает за тем, как экспериментатор берет рукой пищу или другой объект. Однако, тот факт, что зрительный стимул активирует тот же самый нейронный паттерн — то есть, тот же набор потенциальных моторных актов, который отвечает за работу всей моторной цепочки — говорит о том, что обезьяна с самого начала способна ухватить реальную динамику целенаправленных движений, за которыми она наблюдает, и предугадать, чем завершится действие экспериментатора, как только он начнет движение.
Правда, в описываемых экспериментах обезьяне были доступны подсказки, позволяющие ей выбрать подходящее значение для действий, которые она видела, но если бы этих подсказок-индикаторов не было, обезьяне бы потребовалась божественная сила! Контейнер был одной из самых важных подсказок: если он стоял на столе, экспериментатор клал пищу в него, если его не было — то себе в рот. Кроме того, подсказки могли взаимодействовать друг с другом: на самом деле, многие нейроны, отвечающие при наблюдении обезьяной за действием по хватанию пищи и поднесению ее ко рту, разряжались, хотя и более слабо, и при наблюдении за действием по перемещению пищи в другое место (однако, так не происходило при работе с объектами, не являющимися пищей).
Это практически выглядело так, как если бы присутствия пищи в поле зрения и возможности протянуть руку и взять ее было бы достаточно для того, чтобы активировать, пусть и слабо, цепочку действий, которая приведет к тому, что пища будет положена в рот — даже если по контексту будет понятно, что результатом действия будет перенесение пищи в другое место. Ответы других нейронов становятся слабее при повторе действия, как если бы активация цепи «хватания для перемещения» постепенно подавляла активацию цепи «хватания для еды».
Все эти аргументы в целом подтверждают важность моторного знания для понимания действий, в то же время расширяя его роли и функции. Такое знание позволяет нам распознавать значения моторных актов, которые мы наблюдаем — и когда они выполняются вне контекста, и когда они являются частью целой цепи действий. В этом последнем случае их значения более не определяются только лишь специфическими связями между объектами, позволяющими отличать одно действие от другого. Хватание теперь не просто хватание, но хватание для того, чтобы съесть или для того, чтобы переместить. Здесь намерение выполнить действие преобладает над выполнением отдельного движения и изменяет его значение в ту или иную сторону.
Если бы моторные цепочки не были организованы так, как продемонстрировали Фогасси и его коллеги, скорее всего, мозг обезьяны практичеcки не мог бы создавать такие инструкции, которые обеспечивали бы плавность движений и их сходство с тем, что Александр Романович Лурия любил называть «кинетическими мелодиями»18. Более того, без зеркальных свойств этих нейронов обезьяна была бы не способна ни мгновенно уловить, так сказать, намерения, которые наполняли эти мелодии содержанием, когда они проигрывались другими, ни смогла бы также предугадать с самых первых движений промежуточный результат действия (то есть, захват пищи рукой) и, что более важно, его конечный итог — хватание для того, чтобы съесть или чтобы переместить.
Чем более точная информация может быть получена из контекста и от самого объекта, тем более избирательной будет активация соответствующей моторной цепи. Однако, даже если сенсорная информация неопределенна, какой она часто и бывает (и не только в экспериментальных условиях), активация одного или нескольких целенаправленных потенциальных моторных актов помогает обезьяне расшифровать намерения экспериментатора.
После этого животное становится способно выбирать ту интенцию, которая ему кажется наиболее соответствующей сценарию действия, вплоть до распознавания самой подходящей, и использует ее, ничего не говоря о том, что эти процессы расшифровки и идентификации были связаны с тем же моторным знанием, которое направляет и приспосабливает выполнение животным той же самой цепочки действий.
5. Зеркальные нейроны у человека
Первые данные
После обнаружения зеркальных нейронов у обезьян логично было сделать следующий шаг и предположить наличие подобной системы и у человека. Часто (и не только в нейрофизиологии) новые открытия способствуют пересмотру и новой интерпретации старых данных, уже имеющихся в литературе, что и случилось с идеей зеркальных нейронов. Доказательства (часто косвенные), подтверждающие наличие механизма, который мы сейчас интерпретируем как зеркальную систему, были найдены при помощи регистрации ЭЭГ (электроэнцефалограммы) в начале 50-х годов, когда были проведены исследования реактивности ритмов мозга при наблюдении за движением.
Хорошо известно, что при помощи ЭЭГ можно зарегистрировать спонтанную электрическую активность мозга. Ритмы ЭЭГ классифицируются по их частоте. У взрослых здоровых людей в спокойном состоянии при закрытых глазах в задних отделах мозга доминирует альфа-ритм (8—12 Гц), а в передних — так называемые ритмы десинхронизации (имеющие высокую частоту и низкую амплитуду). Еще один ритм, известный как мю-ритм и сходный с альфа-ритмом, регистрируется в центральных областях. Альфа-ритм преобладает при бездействии сенсорных систем, в частности, зрительной. Если зрительные условия меняются и закрытые глаза открываются, этот ритм исчезает или значительно ослабляется. Мю-ритм также отчетливо выражен, когда моторная система находится в покое, а активное движение или соматосенсорная стимуляция десинхронизируют его.
В 1954 году Анри Гасто16 и его коллеги провели эксперименты, в которых показали, что мю-ритм десинхронизируется не только при выполнении действий, но и при наблюдении за действиями других. Спустя сорок с лишним лет, вдохновленные открытием зеркальных нейронов, Вилейанур С. Рамачандран и его коллеги, а также Стефани Кочин и ее коллеги17 воспроизвели эти эксперименты, используя усовершенствованные методы. В частности, группа Кочин показала, что наблюдение за движениями ноги или пальцев сопровождается десинхронизацией мю-ритма, и что этого не происходит, если испытуемым демонстрируется движущийся объект. Другими словами, тот же самый ритм, который блокируется или десинхронизируется движением, также подавляется при наблюдении за движением другого человека.
Похожие результаты были получены в серии исследований, проведенных при помощи МЭГ (магнитоэнцефалографии) — метода, который позволяет анализировать электромагнитную активность мозга, используя регистрацию генерируемых мозгом магнитных полей. Результаты этих исследований также показали, что мю-ритм в прецентральной коре подавляется как при манипуляции с объектами, так и при наблюдении действий других18.
Исследования с использованием транскраниальной магнитной стимуляции (ТМС) также обеспечивают очень убедительные доводы в пользу того, что моторная система человека обладает зеркальной функцией. ТМС — это неинвазивный метод, позволяющий стимулировать мозг. Вплотную к голове подносится электромагнитная катушка, образующееся вокруг которой магнитное поле вызывает индуцированные токи определенной интенсивности в моторной коре. Это позволяет регистрировать моторные потенциалы (двигательные вызванные потенциалы, или ДВП) в мышцах контралатеральной конечности.
Так как амплитуда ДВП модулируется контекстом, в котором выполняется действие, этот метод может быть использован для контроля возбудимости моторной системы в различных экспериментальных условиях. Лучано Фадига и его коллеги19 регистрировали ДВП, вызванные стимуляцией левой моторной коры, в различных мышцах правой руки и кисти испытуемых, которых просили смотреть на то, как экспериментатор берет рукой различные объекты и выполняет движения, не имеющие смысла и никак не связанные с объектом. В обоих случаях было обнаружено избирательное увеличение амплитуд ДВП в мышцах при наблюдении за действием. Несмотря на то, что увеличение амплитуд ДВП во время наблюдения за транзитивными дейcтвиями (связанными с объектами) согласуется с данными, полученными на обезьянах, было весьма неожиданным обнаружить подобный эффект и во время наблюдения за нетранзитивными действиями (не направленными на объект), так как у обезьян зеркальные нейроны не отвечали на наблюдение за не связанными с объектами движениями руки.
Это не единственное различие между зеркальными системами человека и обезьяны. При регистрации ДВП в мышцах руки у здоровых испытуемых, наблюдавших за тем, как экспериментатор выполняет типичные движения захвата, было показано, что активация моторной коры по времени очень точно совпадает с временем выполнения наблюдаемого действия. Судя по этим данным, можно предположить, что зеркальные нейроны у человека способны кодировать не только цель действия, но и временные характеристики отдельных входящих в него движений20.
Данные томографических исследований
Важные функциональные следствия описанных выше результатов будут рассмотрены позднее, сейчас же необходимо представить еще один источник данных, благодаря которому были получены подтверждения существования у человека зеркальной системы — это методы картирования мозга.
Несмотря на то, что электрофизиологические методы, такие как ЭЭГ, МЭГ и ТМС, могут быть использованы для регистрации специфичеcкой активации моторной системы человека, вызванной наблюдением за действиями других, они не позволяют точно локализовать корковые зоны и системы, обеспечивающие такую активность. Следовательно, с их помощью невозможно выяснить строение зеркальной системы в целом. Поэтому необходимо обратиться к методам картирования мозга, в частности, позитронно-эмиссионной томографии (ПЭТ) и функциональной магнитно-резонансной томографии (фМРТ), благодаря которым можно регистрировать вызванные выполнением и наблюдением за определенными моторными актами изменения кровотока в тех или иных областях мозга.
Однако, результаты первых таких исследований были далеки от ожидавшихся21. Участникам эксперимента предъявлялись изображения хватательных движений при помощи руки, созданной в виртуальной реальности. Методом ПЭТ не удалось зарегистрировать какую-либо значимую активацию в моторных зонах, которые соответствуют вентральной премоторной коре у обезьян. Эффекты, обнаруженные в описанных ранее электрофизиологических исследованиях, пока не могли быть объяснены.
Позднее этот эксперимент был воспроизведен другими исследователями22, но с одним важным отличием: движения выполнялись настоящей рукой, а не виртуальной, которая явно была нереалистичной. На этот раз результаты ПЭТ подтвердили то, что было найдено у обезьян: существуют зоны в лобной коре, которые активируются при наблюдении за движениями руки другого. Затем и в фМРТ-исследованиях были более точно локализованы зоны, включенные в зеркальную систему: во время наблюдения за действиями других постоянно активируются ростральная (передняя) часть нижнетеменной коры и нижняя часть прецентральной извилины, а также задняя часть нижнелобной извилины. В определенных экспериментальных условиях также активируются более передняя область нижнелобной извилины и дорзальная премоторная кора (рис. 5.1, см. цв. вклейку).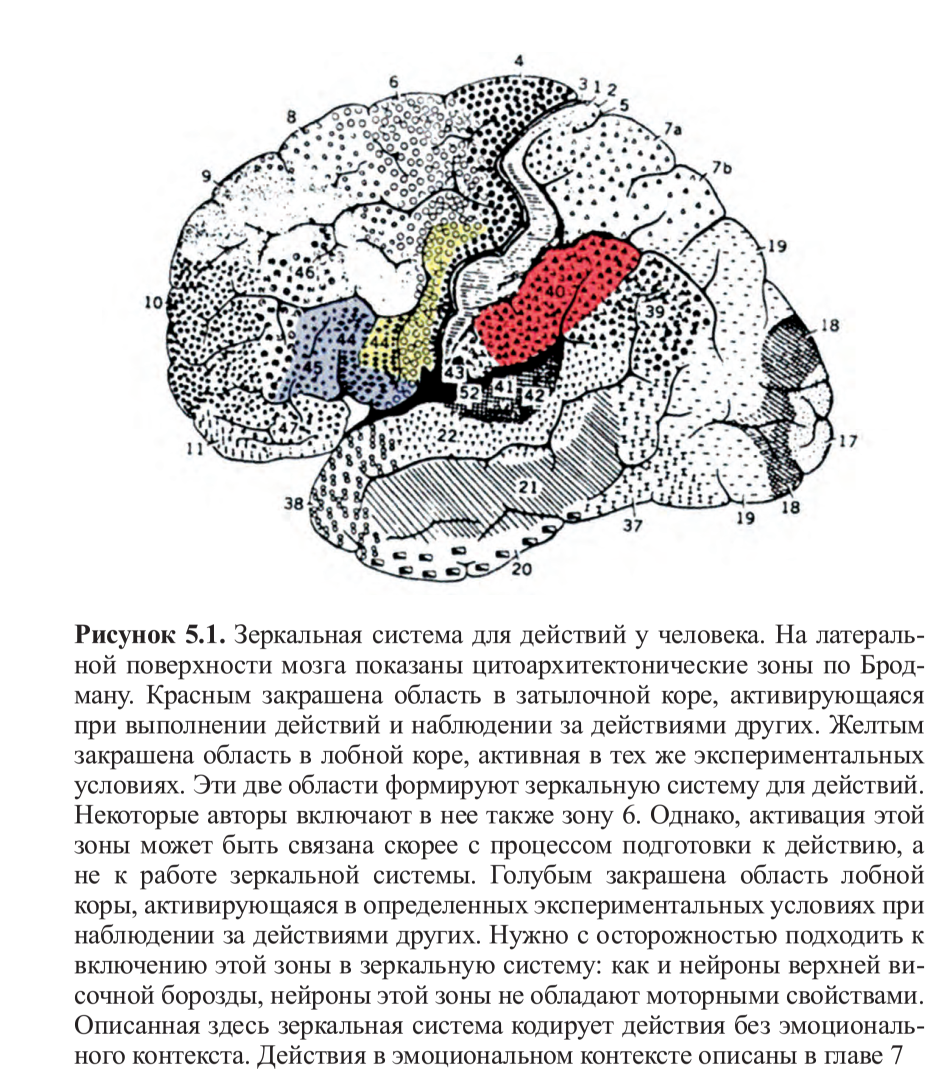 Несмотря на то, что интерпретировать данные о корковой активности, полученные при помощи фМРТ, в терминах цитоархитектонических зон всегда рискованно, очень высока вероятность того, что активирующийся участок нижнетеменной коры соответствует зоне 40 по Бродману — гомологу зоны PF у человека, которая, как мы уже видели ранее, является одной из тех зон, где были найдены зеркальные нейроны у обезьян. Но активацию в нижней части прецентральной извилины и задней части нижнелобной извилины оказалось гораздо сложнее описать цитоархитектонически. Долгие годы считалось, что это две разные зоны, не имеющие никакой функциональной взаимосвязи: задняя часть нижнелобной извилины соответствует зоне 44 по Бродману (то есть, задней части зоны Брока) и отвечает за речь, тогда как нижняя часть прецентральной извилины рассматривается как относящаяся к моторной коре.
Несмотря на то, что интерпретировать данные о корковой активности, полученные при помощи фМРТ, в терминах цитоархитектонических зон всегда рискованно, очень высока вероятность того, что активирующийся участок нижнетеменной коры соответствует зоне 40 по Бродману — гомологу зоны PF у человека, которая, как мы уже видели ранее, является одной из тех зон, где были найдены зеркальные нейроны у обезьян. Но активацию в нижней части прецентральной извилины и задней части нижнелобной извилины оказалось гораздо сложнее описать цитоархитектонически. Долгие годы считалось, что это две разные зоны, не имеющие никакой функциональной взаимосвязи: задняя часть нижнелобной извилины соответствует зоне 44 по Бродману (то есть, задней части зоны Брока) и отвечает за речь, тогда как нижняя часть прецентральной извилины рассматривается как относящаяся к моторной коре.
Нельзя не упомянуть здесь о том, что в самом начале XX века Альфред Уолтер Кэмпбелл, один из отцов цитоархитектоники, обратил внимание на сходство в анатомическом строении задней части нижневисочной извилины и нижней части прецентральной извилины, назвав эту область термином собственного изобретения — «промежуточная прецентральная кора»23. На его наблюдение, однако, никто не обращал внимания в течение многих лет. И только недавние сравнительно-анатомические исследования показали, что зона 44 по Бродману (по крайней мере, ее часть) может рассматриваться как гомолог у человека зоны F5у обезьян24; более того, стало ясно, что в зоне 44 имеются репрезентации руки и рта25.
Может ли теперь, на основании приведенных данных, этой зоне быть приписана ключевая роль в зеркальной нейронной системе человека? Можно ли действительно использовать функциональную гомологичность с зоной F5 у обезьян в качестве ключевого аргумента в пользу того, чтобы интерпретировать полученную в томографических исследованиях активацию как наличие у нейронов задней части нижнелобной извилины свойств, аналогичных зеркальным? Не будет ли проще предположить, что их активация отражает «внутреннюю вербальную репрезентацию»? Мы часто описываем в уме действия, которые наблюдаем — например, мы можем бормотать себе под нос:
«Только вдумайтесь в это — вот, оказывается, как я беру свою чашку кофе!»
Итак, можем ли мы действительно исключать возможность того, что нечто подобное происходит и с испытуемыми в ПЭТ-исследованиях? В конце концов, активируется часть зоны Брока! Джованни Буччино и его коллеги26 попытались ответить на эти возражения, проведя фМРТ-исследование. Они просили нескольких студентов просматривать видеоролики, в которых актеры либо выполняли транзитивные действия, такие как откусывание яблока, поднимание чашки кофе, удар по футбольному мячу, либо изображали их. Наблюдение за транзитивными движениями рта выявило два фокуса активации в лобной коре: один соответствовал задней части нижнелобной извилины, а другой — нижней части прецентральной, и два — в нижнетеменной коре, а наблюдение за транзитивными движениями руки выявило такой же паттерн активации, за исключением того, что активация в нижней части прецентральной извилины была локализована более дорзально, а в ростральной части нижнетеменной коры — более сдвинута назад.
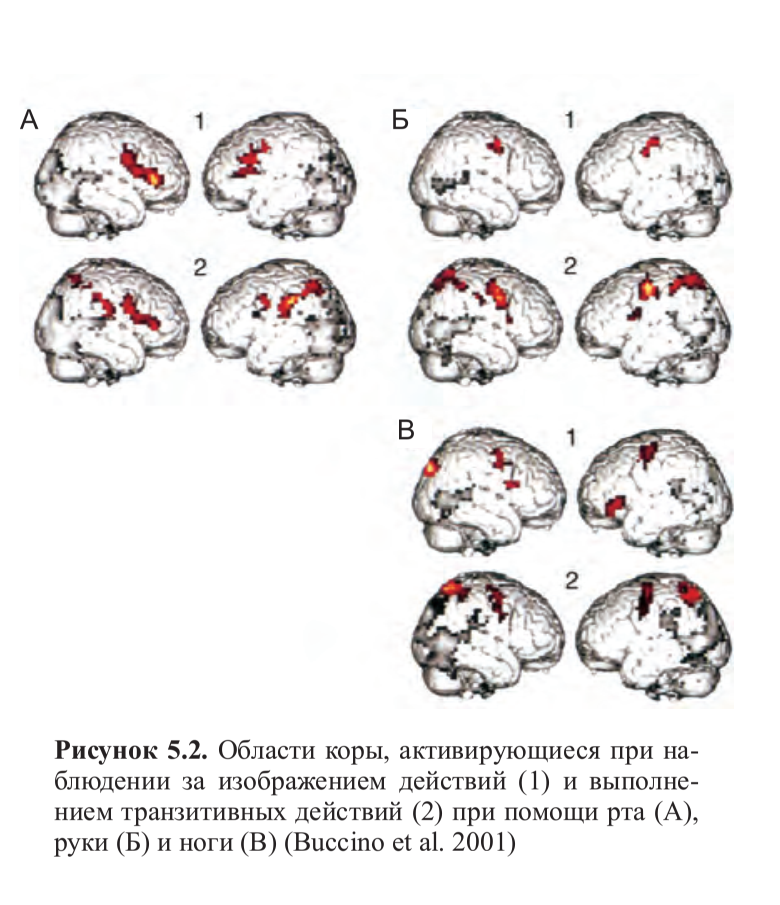
Транзитивные движения ног вызывали фронтальную активацию, расположенную более дорзально по сравнению с активацией на движения руки и рта, и еще больший сдвиг назад фокуса активации в теменной коре. Другими словами, несмотря на значительное перекрытие, зеркальная система, похоже, организована соматотопически и имеет отдельные области для движений рта, рук и ног. При наблюдении за изображением движений был получен аналогичный паттерн активации, ограниченный, однако, только лобной корой (рис. 5.2, см. цв. вклейку).
 Если бы гипотеза о вербальном опосредовании была верна, зона Брока должна была бы активироваться независимо от типа наблюдаемых действий и использованного органа-эффектора, а также не было бы никакой активации премоторной коры. Однако, описанные выше результаты не подтверждают эту гипотезу, и поэтому, если мы не хотим и далее придерживаться довольно причудливого объяснения ad hoc — настаивая, например, на том, что вербальная репрезентация должна присутствовать при наблюдении за движениями рта и руки, но каким-то волшебным образом должна исчезнуть, когда мы смотрим на движения ног, — мы вынуждены признать, что активация зоны Брока отражает типичное поведение зеркальных нейронов. Более того, эксперимент Буччино и коллег показал, что в зеркальную систему человека помимо зоны Брока включены большие области премоторной и нижнетеменной коры. Это еще одно доказательство в пользу того, что зеркальная система отвечает не только на движения руки и транзитивные действия, но и на изображение действий.
Если бы гипотеза о вербальном опосредовании была верна, зона Брока должна была бы активироваться независимо от типа наблюдаемых действий и использованного органа-эффектора, а также не было бы никакой активации премоторной коры. Однако, описанные выше результаты не подтверждают эту гипотезу, и поэтому, если мы не хотим и далее придерживаться довольно причудливого объяснения ad hoc — настаивая, например, на том, что вербальная репрезентация должна присутствовать при наблюдении за движениями рта и руки, но каким-то волшебным образом должна исчезнуть, когда мы смотрим на движения ног, — мы вынуждены признать, что активация зоны Брока отражает типичное поведение зеркальных нейронов. Более того, эксперимент Буччино и коллег показал, что в зеркальную систему человека помимо зоны Брока включены большие области премоторной и нижнетеменной коры. Это еще одно доказательство в пользу того, что зеркальная система отвечает не только на движения руки и транзитивные действия, но и на изображение действий.
Понимание намерений других при помощи зеркальной системы
Мы увидели, что и электрофизиологические, и томографические исследования показали наличие у человека системы, аналогичной зеркальной системе у обезьян. Но между ними есть и существенные различия: у человека зеркальная система занимает большую площадь коры, чем у обезьян. Однако к этому выводу нужно подходить с особой осторожностью и с учетом различий в экспериментальных методах, использованных в исследованиях с обезьянами и человеком: одно дело — регистрировать активность отдельных нейронов, и совсем другое — анализировать активацию различных корковых зон на основании изменений кровотока. Тем не менее, наиболее важно то, что зеркальная система человека обладает некоторыми свойствами, которых не было найдено у обезьян — к примеру, она кодирует как транзитивные, так и нетранзитивные моторные акты, как цели моторных актов, так и движения, из которых состоит законченное действие, и, наконец, в случае с транзитивными действиями, эффективное взаимодействие с объектом не является обязательным условием, так как для активации зеркальной системы достаточно и изображения действия.
Мы уже упоминали, что эти свойства могут иметь важное функциональное значение, однако, тот факт, что зеркальная система человека может выполнять более широкий спектр задач, чем тот, который наблюдался у обезьян, не должен затмевать основную функцию зеркальной системы, связанную с пониманием значения действий других. На самом деле, эксперименты с ТМС показали, что наблюдения за действиями, выполняемыми другими при помощи рук, приводят к увеличению амплитуд ДВП, регистрируемых в мышцах соответтвующей руки наблюдателя, которая задействована в выполнении тех же дейcтвий. С другой стороны, томографические исследования показали, что активация лобной коры при наблюдении за выполнением другими действий при помощи рук, рта и ног определяет активацию, соответствующую соматотопическим моторным репрезентациям этих частей тела.
Как и у обезьян, у людей наблюдение за выполнением действий другими приводит к незамедлительной активации моторных зон, связанных с организацией и выполнением этих действий, и благодаря этой активации становится возможным расшифровать значение наблюдаемых «моторных событий», то есть, понять их в терминах целенаправленных движений. Это понимание полностью лишено каких-либо рефлексивных, концептуальных или лингвистических опосредствований и основано только на словаре действий и моторном знании, от которого зависит наша способность действовать. Наконец, как и у обезьян, это понимание не ограничено отдельными моторными актами, но может распространяться на целые их цепочки.
Это последнее их свойство очень явно проявляется в фМРТ-исследованиях, проведенных Марко Якобони и его коллегами27. Они показывали добровольцам три различных видеоклипа (рис. 5.3). На первом клипе участники эксперимента видели ряд объектов (чайник, кружку, стакан, тарелку, и т. д.), которые лежали на столе так, как будто кто- то собирался пить чай (а не кофе из нашей привычной чашки!) или только что его пил. Экспериментальное условие, в котором предъявлялось это видео, для краткости было названо «контекст». На втором видеоклипе была показана рука, захватывающая кружку при помощи всей кисти либо точного захвата при отсутствии контекста (условие «действие»). На третьем клипе была та же самая рука, согнутая тем же самым образом, но на этот раз в контексте, который позволил бы предположить, что у действующего есть намерение взять чашку со стола либо для того, чтобы поднести ее к губам, либо чтобы убрать ее со стола после чаепития (условие было названо «намерение»).

Рисунок 5.3. Видеоклипы, использованные в исследовании корковых зон, включенных в понимание намерений других. Клипы, соответствующие разным экспериментальным условиям, расположены в три колонки: контекст, действие, намерение. В условии контекст (первая колонка) участники эксперимента видели стол, накрытый для завтрака (вверху) и тот же стол после завтрака (внизу). В условии действие (вторая колонка) они видели руку, которая брала чашку со стола, используя при этом либо захват всей кистью (вверху), либо точный захват (внизу). Контекста в этом условии не было.
В третьем условии (намерение) были показаны два типа захвата в контексте «до» и «после» завтрака, предполагая, соответственно, намерения действующего «взять кружку, чтобы пить из нее» (вверху) и «взять кружку, чтобы помыть ее» (внизу) (Iacoboni et al. 2005).
Если мы сравним активность мозга, вызванную наблюдением действий в трех различных условиях, с контрольным условием отдых (рис. 5.4, см. цв. вклейку), мы увидим, что в условиях действие и намерение активность увеличивалась в зрительных зонах и в лобнотеменной системе, связанной с кодированием моторных актов, тогда как в условии контекст не было увеличения активности ни в области STS (верхняя височная борозда), отвечающей на движение зрительных стимулов, ни в нижнетеменной коре, хотя активность в премоторных зонах была выражена отчетливо. Это можно объяснить присутствием объектов, которые можно схватить и которые активируют канонические нейроны, отвечающие, как мы видели, на возможности, предоставляемые объектами.
Наибольший интерес представляет сравнение условий намерение и действие, а также намерение и контекст. Как можно увидеть на рисунке 5.5 (см. цв. вклейку), активация дорзальной поверхности задней части правой нижней лобной извилины (рис. 5.5, вверху) была выше в условии намерение, чем в двух других условиях (действие и контекст). Это особенно интересно, потому что фокус активации расположен в лобной части зеркальной системы — это говорит о том, что зеркальные нейроны не только кодируют наблюдаемое действие (в данном случае, хватание чашки при помощи определенного типа захвата), но также и намерение, с которым данное действие выполняется. Это может быть объяснено тем, что в то время как наблюдатель смотрит, как другой выполняет действие, он уже предугадывает возможные следующие действия в цепочке (к примеру, «взять, чтобы выпить чаю» или «взять, чтобы переставить»).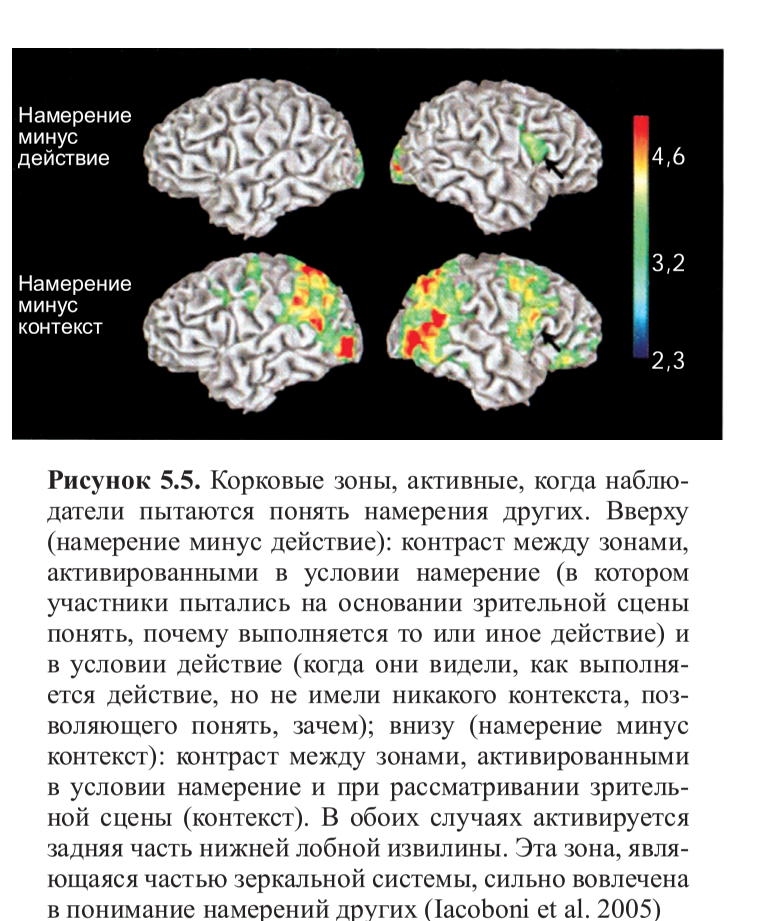 Интересно отметить, что наблюдение за действием поднесения ко рту вызывает более сильную активацию в нейронах зеркальной системы, чем наблюдение за захватом чашки, чтобы убрать ее со стола (рис. 5.6). Это согласуется с данными, полученными Фогасси и коллегами и описанными в предыдущей главе. В их эксперименте было показано, что количество нейронов, кодирующих действие «захвата для еды», больше, чем кодирующих «захват для перекладывания». Кроме того, даже когда сенсорная информация относительно контекста (присутствие контейнера для пищи) предполагает, что наиболее вероятным действием после захвата пищи будет перекладывание ее в контейнер, наблюдение за рукой экспериментатора, берущей пищу, вызывает активность, пусть и достаточно слабую, в той цепочке нейронов, которая отвечает за кодирование «хватания для еды», так как это естественная последовательность действий в репертуаре обезьяны. В исследовании Якобони были получены похожие результаты: активация правой нижнелобной коры была больше при наблюдении за действием «поднести кружку ко рту, чтобы пить из нее», чем в случае «хватания кружки, чтобы помыть ее». Всегда преобладает именно самое естественное двигательное намерение, наиболее глубоко укоренившееся в нашем словаре действий.
Интересно отметить, что наблюдение за действием поднесения ко рту вызывает более сильную активацию в нейронах зеркальной системы, чем наблюдение за захватом чашки, чтобы убрать ее со стола (рис. 5.6). Это согласуется с данными, полученными Фогасси и коллегами и описанными в предыдущей главе. В их эксперименте было показано, что количество нейронов, кодирующих действие «захвата для еды», больше, чем кодирующих «захват для перекладывания». Кроме того, даже когда сенсорная информация относительно контекста (присутствие контейнера для пищи) предполагает, что наиболее вероятным действием после захвата пищи будет перекладывание ее в контейнер, наблюдение за рукой экспериментатора, берущей пищу, вызывает активность, пусть и достаточно слабую, в той цепочке нейронов, которая отвечает за кодирование «хватания для еды», так как это естественная последовательность действий в репертуаре обезьяны. В исследовании Якобони были получены похожие результаты: активация правой нижнелобной коры была больше при наблюдении за действием «поднести кружку ко рту, чтобы пить из нее», чем в случае «хватания кружки, чтобы помыть ее». Всегда преобладает именно самое естественное двигательное намерение, наиболее глубоко укоренившееся в нашем словаре действий.
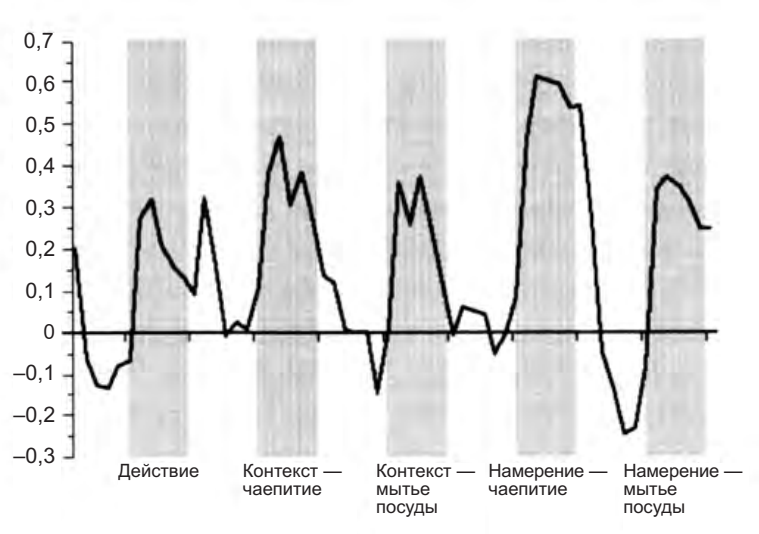
Рисунок 5.6. Временная развертка активности правой нижнелобной коры демонстрирует увеличение сигнала при сопоставлении условий (контрасты намерение минус действие и намерение минус контекст). Как показано на предыдущем рисунке, это зона, активирующаяся, когда наблюдатель «прочитывает» намерения других. Интересно отметить, что наблюдение действия «взять, чтобы выпить чаю» порождает более сильную активацию зеркальной системы, чем за действием «взять, чтобы помыть» (Iacoboni et al. 2005)
Не стоит и говорить, что, как и в случае с моторными актами, активация зеркальных нейронов — не единственное средство, благодаря которому мы способны понимать намерения, заложенные в действиях других. Каждый день мы приписываем другим — более или менее осознанно — убеждения, желания, ожидания, намерения, и т. д. Наше собственное социальное поведение в основном зависит от нашей способности понимать, что другие имеют в виду, и руководствуясь этим, мы решаем, как нам подстроиться под собеседника.
В настоящее время не найден еще такой нейронный механизм, который объяснил бы чтение мыслей: может случиться так, что он окажется эволюционно связанным с зеркальной системой. Что, однако, имеет значение — то, что зеркальный механизм фиксирует интенциональный аспект действий, общий для наблюдателя и исполнителя. Здесь будет очень к месту цитата из Мерло-Понти:
“Смысл жестов не дан, он понимается, то есть улавливается посредством действия наблюдателя. Вся трудность в том, чтобы правильно понять это действие и не спутать его с познавательной операцией. Общение, или понимание жестов, достигается во взаимности моих интенций и жестов другого, моих жестов и интенций, читающихся в поведении другого. Все происходит так, как если бы интенции другого населяли мое тело, а мои интенции населяли тело другого. Жест, свидетелем которого я являюсь, «очерчивает пунктиром» интенциональный объект. Этот объект становится актуальным и полностью понимается, когда способности моего тела приспосабливаются к нему и его охватывают28.
«Действия наблюдателя» — это потенциальный моторный акт, определяемый активацией зеркальных нейронов, кодирующих сенсорную информацию в моторных терминах, таким образом обеспечивая «реципрокность» действий и намерений, лежащую в основе нашей способности мгновенно понимать, что делают другие. Понимание намерений других не основано, в таком случае, на осмыслении, то есть, на метарепрезентации, но зависит от выбора наиболее подходящей к данной ситуации возможной цепочки действий.
Как только мы видим, как кто-то делает что-либо — отдельное действие или цепочку действий — его движения незамедлительно приобретают для нас смысл, хочет он этого или нет. И, очевидно, верно и обратное: наши действия незамедлительно обретают смысл для тех, кто их видит. Зеркальная система и такая избирательность ответов составляющих ее нейронов способствуют созданию общего пространства действий (shared space of actions), внутри которого каждое действие и цепочка действий, принадлежат они «нам» или «другим», мгновенно регистрируются и распознаются без необходимости какой-либо эксплицитной или намеренной «когнитивной операции».
Различия в словарях
Что произойдет, если окажется, что в нашем словаре действий нет того движения, которое мы наблюдаем? В экспериментах, рассмотренных в предыдущей главе, мы видели, что у обезьян зеркальные нейроны разряжаются не только когда другая обезьяна хватает пищу, но также когда действие выполняет экспериментатор. Это не удивительно, если вспомнить, что хватание присутствует в словаре действий обезьяны (равно как и удержание, разрывание, толкание, бросание, и т. д.). И тем не менее, мы часто видим действия, не являющиеся частью нашего моторного знания — возможно, они не входят в репертуар нашего вида, или же просто мы не способны их выполнять.

Рисунок 5.7. Стимульный материал, использованный в эксперименте, целью которого была регистрация активности мозга при наблюдении за действиями, общими для человека и животных. Кадры из видеоклипов показывают, как человек, обезьяна и собака откусывают кусочки пищи (Buccino et al. 2004a)
В одном недавнем исследовании с использованием фМРТ29 группе добровольцев предъявлялись видеоклипы без звука, в которых представители разных биологических видов (человек, обезьяна, собака) выполняли действия, связанные с употреблением пищи (откусывание) или коммуникативные акты (говорение, чмокание губами, лай).

Рисунок 5.8. Стимульный материал, использованный в эксперименте, в котором сравнивалось восприятие коммуникативных жестов человека и представителей других видов. Кадры из видеоклипов показывают человека, обезьяну и собаку в процессе говорения, чмокания губами и лая, соответственно (Buccino et al. 2004a)
На рисунках 5.7 и 5.8 представлены отдельные кадры из этих клипов. Несмотря на то, что вид человека, откусывающего кусочек пищи, сильно отличается от вида обезьяны или (еще больше) от вида собаки, было обнаружено значительное перекрытие зон активности во всех трех случаях. На самом деле, просмотр всех трех видеоклипов порождал два фокуса активности (передний и задний) в нижнетеменной коре, а также в задней части нижней лобной извилины и примыкающей к ней прецентральной извилины. При сравнении трех условий наблюдалась определенная асимметрия между левым и правым полушариями: активация в левом полушарии практически не зависела от того, кем выполнялось действие — человеком, обезьяной или собакой, тогда как в правом полушарии, напротив, активация была сильнее при наблюдении за выполняющим действия человеком (рис. 5.9, см. цв. вклейку).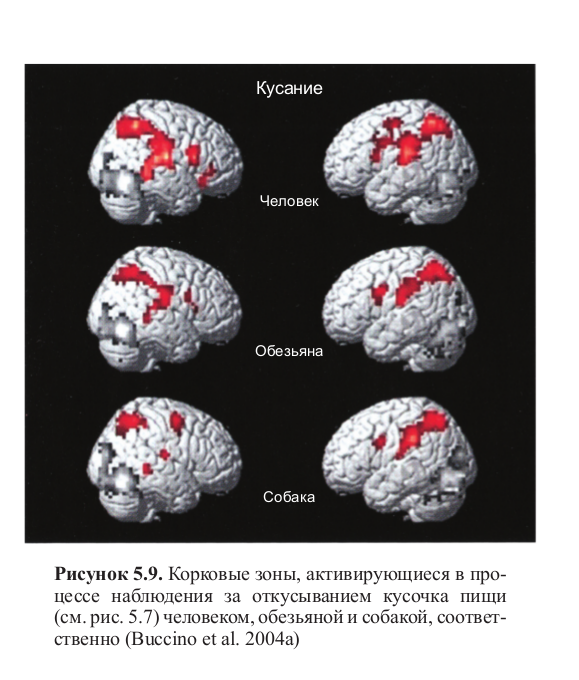
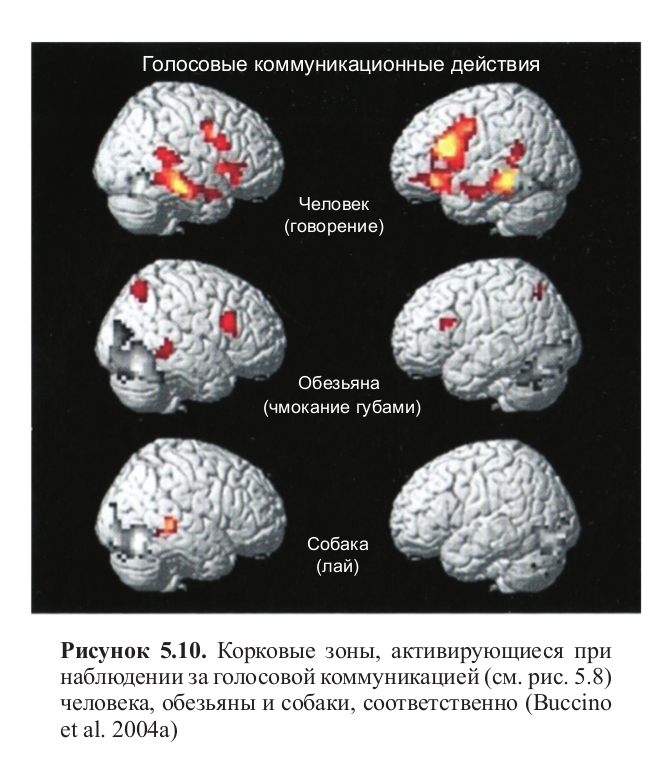 При наблюдении за коммуникативными действиями паттерн активации был совершенно иным. Наблюдение за человеком, двигающим губами так, как будто он разговаривает, вызывало сильную активацию в задней части нижней лобной извилины (область, соответствующая зоне Брока); активация становилась слабее, когда испытуемые наблюдали за чмокающей губами обезьяной, и исчезала совсем, когда они видели лающую собаку (рис. 5.10, см. цв. вклейку).
При наблюдении за коммуникативными действиями паттерн активации был совершенно иным. Наблюдение за человеком, двигающим губами так, как будто он разговаривает, вызывало сильную активацию в задней части нижней лобной извилины (область, соответствующая зоне Брока); активация становилась слабее, когда испытуемые наблюдали за чмокающей губами обезьяной, и исчезала совсем, когда они видели лающую собаку (рис. 5.10, см. цв. вклейку).
В терминах чисто зрительного восприятия различие между коммуникативными актами не окажется большим, чем между действием по откусыванию пищи, выполняемым тремя представителями разных видов, и, следовательно, не сможет объяснить полученные паттерны активации. Отсутствие активности в зеркальной системе при восприятии лающей собаки не может быть просто объяснено типом получаемой зрительной информации. В предыдущей главе мы увидели, что активность зеркальных нейронов не привязана к какой-либо одной модальности — она ограничена словарем действий, регулирующим организацию и выполнение движений. Лай просто не принадлежит к словарю действий человека.
Означает ли это, что мы неспособны понимать движения лающей собаки и отличать их от того, как она откусывает кусочек пищи? Конечно, нет! Просто восприятие этих двух процессов зависит от двух совершенно различных аспектов понимания: первый преимущественно основывается на зрительной информации, а второй — на зрительно-моторной. Когда мы видим лающую собаку, наше понимание ее действий связано преимущественно с активацией зон верхней височной борозды (STS). Эти, а также другие зрительные зоны активируются при наблюдении за коммуникативными действиями других (людей или обезьян), но в этом последнем случае информация, поступающая из STS, запускает потенциальный моторный акт, кодирующийся в зеркальной системе, благодаря чему мы незамедлительно понимаем значение воспринимаемых нами действий30.
Подобные различия были найдены и между индивидами одного вида. Беатрис Кальво-Мерино и ее коллеги31 провели фМРТ-исследование, в котором было показано, что корковая активность может меняться в зависимости от наличия специфических моторных навыков наблюдателей за выполнением этих специфических действий другими. Испытуемыми были классические танцовщики, тренеры капоэйры и люди, никогда не занимавшиеся танцами. Им показывали видеоклипы урока капоэйры и классических балетных па. Зеркальные системы у тренеров активировались сильнее при просмотре записи капоэйры, а зеркальные системы танцовщиков — при просмотре балетных па, в отличие от тренеров и, тем более, новичков.
В дальнейших экспери
Свежие комментарии