 Print
Print  PDF
PDF
Горная гаичка и самец пушистопёрого дятла на кормушке
Резюме. Обсуждается непригодность модели доместикации Д.К.Беляева для понимания изменений, происходящих в экологическом и эволюционном масштабах времени при освоении городов «дикими» видами птиц и млекопитающих. Доместикация и урбанизация часто отождествляются, т.к. оба считаются «приспособлением к человеку и созданной им искусственной среде обитания», т.е. развитие синантропности. Проведённый анализ показывает ошибочность этого отождествления. При урбанизации «диких» видов мозг увеличивается, как и в других вариантах экстремальной среды обитания. Когнитивный прогресс достигается каждой «городской» особью самостоятельно, за счёт развивающего воздействия урбосреды, в оценивании и прогнозе динамики урбосреды по сигналам. Поэтому он предваряется подъёмом смелостью, лучшей дифференциацией стимулов, отделением значимых (они отреагируются) от всех остальных, к которым растёт резистентность. Напротив, при одомашнивании мозг уменьшается, когнитивный прогресс в новой среде обитания достигнут за счёт «кооперативности мышления», социальных «подсказок» людей и сородичей. Демонстрируется сомнительность гипотезы «самоодомашнивания» бонобо, развитой в рамках этого же отождествления, и частичная применимость её к человеку.
Главный фактор, к которому приспосабливаются при урбанизации (в меньшей выраженности — в заселении других антропогенно изменённых ландшафтов) — средовой стресс, связанный с общей нестабильностью и изменчивостью урбосреды, в т.ч. кружева местообитаний вида в городе, особенно, когда это «осколки» природных ландшафтов, захваченные при территориальном росте городов и более-менее изменённые за время существования внутри городских ареалов.
Постановка проблемы
Содержание
В статье сопоставлены поведенческие изменения, с одной стороны, в «городских» популяциях птиц сравнительно с «сельскими» или «лесными», с другой — одомашненных видов в среде, созданной для них человеком. Они часто отождествляются, их многочисленные различия игнорируются, почему и заслуживают специального внимания. Один из авторов грешен, сам так делал на основе сходных изменений гормонального уровня в обоих, и по той же причине до 2014 г. уподоблял урбанизацию и доместикацию, не обращая внимание на различия.
Так или иначе, по завершении обоих процессов содержание кортикостерона в целом снижается, а серотонина увеличивается (Bonier, 2012; Deviche, Davies, 2014; Partecke, 2014). Чем дальше зашла урбанизация вида, тем меньше первоначально чужеродная среда и её составляющие стрессируют птиц, всё реже запускают тревожно-оборонительную реакцию и чаще — исследование, манипуляцию и игру как формы реакции на новизну, созданную постоянными изменениями обстановки вокруг: «кружева» местообитаний, обеспеченности их ресурсами, беспокоящих контактов с людьми и пр. Здесь более сложная и изменчивая урбосреда (сравнительно с местообитанием исходной популяции) оказывает развивающее воздействие вместо прежнего подавления.
Когнитивные «достижения» урбанизированных популяций
У видов, где это специально исследовали, она увеличивает когнитивные «достижения» «городских» популяций сравнительно с исходными
I) по сложности задач, которые они способны решать, естественно возникающих или поставленных экспериментатором. Среди них — инновации, проблемные задачи, лучшее привыкание к беспокойству со стороны человека, лёгкость обучения и переучивания, качество долговременной памяти, иногда — изменения объёма переднего мозга. См.обзоры Miranda, 2017, table.7.1; Sol et al., 2020, table 15.1 и анализ Sol et al., 2013, 2016; Dukatez et al., 2017, 2020;
II) по смелости — мотивации их решать, оказавшись в новой и необычной среде и, шире, вообще направленно действовать действовать в этих условиях, достигая решения в т.ч. путём проб и ошибок, вместо неофобии, страха и отступления, наиболее обычных в исходных популяциях (Lowry et al., 2013; Miranda, 2017; Sepp et al., 2020);
III) по доле особей, демонстрирующий высший уровень интеллекта и смелости: в «городских» популяциях их большинство или почти все, в «сельских» или «лесных» — только немногие, в первую очередь «лучшие» особи, выигрывающие конкуренцию за территорию и партнёра, доминанты жизни в стаях и пр. (Фридман, 2018; Фридман, Суслов, 2018).
IV) Разрушение поведенческого синдрома, определяющего реакцию особей на новую и неизвестную ситуацию, беспокойство пешеходами, собаками, велосипедистами и т. д. объектами, прямо не определяемыми как «хищники», но также потенциально опасными (англ. Behavioural syndrome, Sih et al., 2004; Sih, de Guidice, 2012). Составляющие его разные виды активности до урбанизации жёстко связаны между собой корреляциями, положительными и отрицательными, первые проявляются вместе, дополняя или усиливая поведенческий ответ, вторые исключают друг друга. Когда ситуация вызывает беспокойство и стресс, запускается оборонительное поведение, составляющие которого связаны с отдельными моментами развития ситуации: сперва отслеживание объекта, потом общий рост настороженности и страха (учащение осматриваний, замирание и пр.), сперва чередующийся с попытками кормиться, потом бегство (на одноименной дистанции, англ. flight distance1).
Они коррелируют отрицательно с «выдачей» поведения, связанного с исследованием и смелостью, позволяющим проявить интеллект и воспользоваться случившимся в своих интересах, исключив неофобию и другие реакции, подавляющие всё названное (Atwell et al., 2012; Miranda et al., 2013; Carrete, Tella, 2017; Dukatez et al., 2017). Т.е. альтернативы реагирования на новизну в исходных популяциях соответствуют имеющимся у доместицированных vs агрессивных лисиц, лабораторных грызунов и т. д. (Hare et al., 2012). Однако урбанизация, в противоположность доместикации, отнюдь не ведёт к смене первого набора корреляций вторым. В «городских» популяциях названные корреляции разрушаются: каждая составляющая в исследовании ситуации, манипуляции и игре с образующими её объектами или, наоборот, в действиях тревоги и бегства проявляется независимо от прочих, «запускаясь» собственными изменениями в урбосреде.

Стенная ящерица
«Рассыпавшись» поодиночке, формы поведения ранее единого синдрома позволяют «городским» особям в каждой ситуации выбирать, как определить данную ситуацию, «опасность», «безразличие» или «интерес» и «возможная польза», какие её составляющие отреагировать и как. Поэтому исследование, рассудочная деятельность, активное использование людей и техники сплошь и рядом сопряжено с сохраняющейся неофобией, немедленным бегством когда на птиц вдруг обращают внимание, пытаются их снимать, приближаться и пр. Реже урбанизация перестраивает прежний синдром в качественно иной; см. анализ разрушения поведенческого синдрома при урбанизации разных видов птиц.
Успешная урбанизация «приспосабливает» особей к собственному стрессу, вызванному островным эффектом, необходимостью перемещения с одного «острова» местообитаний на другой, беспокойством людьми и техникой, другими факторами, значимыми для исходных популяций. В ходе урбанизации этот средовой по происхождению стресс отнюдь не уменьшается, однако становится всё менее травматичен для «городских» птиц, в чём и заключается их адаптация к урбосреде. В такой ситуации её сложность, гетерогенность и вариативность составляющих оказывают развивающее воздействие, вместо прежнего подавления (Мешкова, Федорович, 1996: 43-46): продлевают период обучения, поддерживают исследования нового и непонятного вместо беспокойства и бегства, стимулируют манипуляцию и игру (Sol et al., 2013, table 1; Preiszner et al., 2016; Sol et al., 2020, table 15.1), включая «прогнозы» её динамики по сигналам-предвестникам (Фридман, Ерёмкин, 2009: 61-63; Lowry et al., 2013; Griffin et al., 2017).
Здесь важнее всего данные Н.Н.Мешковой и Е.Ю. Фёдорович (1996: 78-86) о повелении городских домовых мышей в «жилой комнате». Они интенсивно исследуют компоненты новой и сложной среды, играют с ними, а дальше используют в собственном поведении, скажем, для ухода от агрессии доминантов. Сходным образом группы «городских» домовых воробьёв решали проблемные задачи быстрее «сельских», т. к. в ответ на их предъявление превалировала готовность исследовать, а не страх (Liker, Bókony, 2009). Негородские мыши, наоборот, сложной среды избегают из-за стрессирования. То же мы видим у склонной к урбанизации восточноевропейской полёвки Microtus rossiaemeridionalis в отличие от серой M.arvalis, урбанизирующейся гораздо труднее (Тихонова и др., 2012: 284-300).
Эти различия сохраняются и при выкармливании детёнышей человеком, т. е. представляют собой продукты изменений на генетическом уровне, поддержанных отбором. Опытами с выращиванием в изоляции также показан врождённый характер изменений у «городских» серых юнко2 (большая смелость, меньшая неофобия, исследование нового пространства происходит быстрее, но менее тщательно, его отрицательная корреляция с максимумом выброса кортикостерона в ответ на стрессор, но не с базовым уровнем или средним подъёмом гормона в подобном случае, при сниженности того и другого показателя3, Atwell et al., 2012); черных дроздов (по степени неофилии — сближения с объектом в знакомой обстановке и неофобии — его избегания в незнакомой, Miranda et al., 2013); горных гаичек Parus gambeli, освоивших г.Рино (Невада, США). «Городские» гаички лучше справляются с новыми проблемными задачами, быстрее и интенсивней обследуют новую среду, лучше удерживают пространственную информацию в долговременной памяти. Не найдено значимых отличий от «лесных» птиц в уровне неофобии, интенсивности запасания пищи, объёме и общем количестве нейронов гиппокампа (Kozlovsky et al., 2017).

Черноголовая гаичка
Сходный результат дали опыты с искусственным выращиванием водяных легуанов Intellagama lesueurii из природных и пригородных (сады и парки) vs городских ландшафтов. Пять раз за первый год жизни у них оценивали такие черты индивидуальности, как смелость, неофилия, исследовательская активность, определяя их выраженность и повторяемость (Baxter-Gilbert et al., fig.1, table 2). Значимые межгрупповые различия отмечены только по смелости: у «пригородных» ящериц она несколько выше, чем у «городских», у тех и других — значительно выше, чем в исходных биотопах. Отмечена тенденция к усилению исследовательской активности по мере урбанизации. Значимая индивидуальная повторяемость отмечена только для смелости, однако у «городских» ящериц её уровень вариативней, чем у «пригородных» (Baxter-Gilbert et al., 2019). Скальные агамы Psammophilus dorsalis из пригородов Бенгалора (Индия) быстрее выучивают места расположения убежищ, и быстрей переучиваются при изменениях сравнительно с «сельскими» (Batabyal, Thaker, 2019).

Перестройки внутри города способствуют заселению задних дворов Сиднея водяным легуаном
Аналогичные изменения описаны у ряда других видов. «Городские» стенные ящерицы Podarсis muralis смелей: меньше времени проводят в укрытии при угрозе нападения хищника и быстрей привыкают к ней. При последовательных имитациях нападения их время пребывания там снижается сильней, чем у «сельских» (Pelliteri-Rosa et al., 2017). Есть и исключения: успешно заселившие города Восточной Австралии сцинки Lampropholis delicata не отличаются от «сельских» ящериц ни в восприятии риска (реакция избегания новых предметов, запахов или звуков), ни в пространственной памяти (скорость решения стандартного теста на выбор между безопасным vs уязвимым убежищем в Y-образным лабиринте). Возможная причина этого — даже у «сельских» ящериц оба параметра и так максимальны сравнительно с прочими видами, урбанизации «некуда» их увеличить (Walsh et al., 2018).
При большей смелости у «городских» A.sagrei во Флориде усилена исследовательская активность сравнительно с ближайшей «лесной» популяцией; у разных видов Anolis урбанизация также увеличивает осторожность при кормлении. К людям, наоборот, они доверчивей, дистанция вспугивания «городских» ящериц падает также как у птиц. Они лучше дифференцируют реальную опасность от «просто беспокойства» (Lapiedra, 2018).
«Городское» повышение риска, от хищничества до повышенной гибели на дорожках и автотрассах, меняет поведение ящериц в сторону, противоположную природным ландшафтам с повышенным риском хищничества.
В опытах с интродукцией A.sagrei на малые карибские острова с наличием и отсутствием наземного хищника (килеватой масковой игуаны Leiocephalus carinatus) показано, что в первом случае отбор по поведению жёстче, чем по морфологии. Он благоприятствовал более боязливым ящерицам (склонным отступать при потенциальной опасности вместо принятия риска, дольше отсиживающимся в укрытии), и меньше исследующим территорию. В отсутствие хищников, наоборот, выживаемость обратно зависит от длины латентного периода между концом опасности и выходом из укрытия для обследования пространства (Lapiedra et al., 2018).
Сходным образом становились смелей далматинские ящерицы P.melisellensis, освоившие города на более крупных островах Адриатики (о.Вис) сравнительно с островами поменьше, лишь с сельскими поселениями (о. Biševo и Veli Budikovac), и вовсе незаселёнными, без естественных хищников (о.Brusnik). Принятие риска исследовали, предъявляя модели потенциальной опасности — змеи и грача Corvus frugilegus, вызывающие бросок в укрытия и более или менее долгое пребывание там, прежде чем ящерица возобновит кормление. Его длительность — мера страха перед опасностью или, наоборот, готовности к принятию риска (больше всего ящериц пугала змея). Одновременно их тестировали на неофобию, предъявляя новый, возможно пугающий, но полезный объект — ярко-красную пластиковую бутылку с водой, из которой ящерицы могли пить (если не боятся её) (de Meestr et al., 2018).
Ящерицы на обитаемых островах быcтрее высовывали голову из укрытия после предъявления модели хищника, но с меньшей вероятностью подходили к кормушке с мучными червями несмотря на рядом находящуюся модель змеи, при большей длительности задержки у подходивших. На необитаемых островках большая доля ящериц кормилась в таких условиях. Отдельных ящериц можно было кормить с рук, они бегали по ним, облизывали и кусали. Разницы в восприятии опасности «городскими» и «сельскими» ящерицами не найдено, возможно из-за невысокой степени урбанизации. Всем популяциям ящериц неофобия была вовсе несвойственна, предъявление красной бутылки отнюдь не препятствовало подходу к кормушке. На о.Брусник, однако, часть особей демонстрировали неофилию: бутылка их привлекала настолько, что они её обследовали в ущерб кормлению (de Meestr et al., 2018).
Т.е. урбанизация «заставляет» ящериц в крупных городах большую часть времени вести себя так, как в природных ландшафтах с отсутствием хищника, в то же время непрерывно отслеживая обстановку вокруг, чтобы с опережением реагировать на опасность в те немногие промежутки времени, когда это необходимо. Здесь невозможно не видеть «развивающего» влияния урбосреды.

Скальная агама
Самые яркие примеры изменений I-IV показывают птицы и млекопитающие. Черноклювые сороки Pica hudsonica часто гнездятся поблизости от американских ворон Corvus brachyrhunchos, в т.ч. потому что их агрессивность спасает от разорения гнёзд другими воронами. В Jackson Hole, Вайоминг, исследовалось, как тогда обстоят дела с конкуренцией: «платят» сороки «штраф» за обитание на участке более крупного и агрессивного вида, способного отнять корм у мелкого и/или быстрее добыть часть его, или «соображают», как избежать «штрафа»? В опытах Rhea J.Esposito (2015) использовались т. н. Cheeto — чипсы со вкусом сыра ярко-оранжевого цвета, любимые обоими видами. Их выкладывали у гнёзд ворон и сорок, анализируя, кто выиграет «соревнование» за лакомство, и как быстро.

Черноклювая сорока в Jackson Hole, Вайоминг
Считается, что сороки уступают воронам по интеллекту, однако они быстрей замечают и схватывают чипсы (в среднем на 20 с). Вороны более неофобны и дольше осторожничают, но, определив Cheeto как пищу, они их таскают ловчее. Гнездовые пары обоих видов быстро выучивают, что чипсы могут быть как у их собственного гнезда, так и у чужого, так что можно «брать своё» или «воровать». Вороны таскают чипсы от сорочьих гнезд в 3 раза чаще, чем сороки от вороньих.

Экспериментальная установка с чипсами
Тогда увеличили сложность опыта, заложив чипсы внутрь полой колоды: чтобы их взять, птицы тянули за верёвочку, т. е. сколько-то времени приложенные усилия были безрезультатны. Сороки и здесь меньше боялись задачи, смелей решали головоломку, и добывали свой чипс почти на минуту быстрее ворон. Дальше его помещали на равном расстоянии от гнезда обоих видов, анализируя, кто быстрей схватит чипс. В отличие от выкладывания на ничейную территорию, здесь оба вида одинаково быстро определяли его как лакомство. Однако потом вороны «использовали» поведение сорок как знак наличия корма, ждали, когда сорока спустится и попробует взять чипс, затем спугивали её и хватали сами. Такие сороки-жертвы клептопаразитизма со временем отказываются не только брать что-либо, но даже спускаться на землю (Esposito, 2015).
[embedded content]
Возможно, вороны успешнее в воровстве, а сороки легче им уступают, поскольку меньше боятся нападения хищников на земле: спустившиеся сороки им «показали», что корм безопасен. Объяснение, что они «просто сильнее», менее вероятно, тогда они бы имели преимущество и на «ничейной территории». R.J.Esposito (2015) показывает, что сороки, гнездящиеся рядом с воронами, действительно, защищены от хищничества, но общий успех размножения не возрастает. Возможно, потому что «плата» от уменьшения поступления корма из-за клептопаразитизма сравнима с «выигрышем». В противоположность обычным адаптационистским объяснениям урбанизации, «городские» сороки не «конвертируют» большую смелость в прибавку ресурсов, а меньшая — не мешает воронам урбанизироваться в той же степени (Esposito, 2015).
Важнее всего, что здесь исследовали не интеллект: задачи простейшие, врановые решают куда более сложные, скажем, протоорудийные (Обозова и др., 2013) или на добывание приманки из трубки с ловушкой (Багоцкая и др., 2013). Фактически изучали «сообразительность» — способность
1) использовать новую cитуацию и/или необычный объект в своих интересах, вместо того чтоб пугаться, и
2) быстро переключаться между разными ситуациями, возникающими в одном и том же пространстве-времени активности особи, но требующими противоположных реакций в зависимости от контекста, т. е. его быстрого различения.
Это и отличает «городских» сорок и ворон. Другой важный вывод из этого и других аналогичных исследований: в городских популяциях более смелые и «сообразительные» одновременно (=быстрее воспользовавшихся выгодным изменением в крайне гетерогенном окружении) выигрывают у «более умных» per se, и урбосреда развивает первое сильнее второго. Точней, интеллект прогрессирует вслед за смелостью с некоторым запозданием, т. к. только она создаёт предпосылки для его проявления и совершенствования в актах использования (Dukatez et al., 2020; Sol et al., 2020).

Американская ворона с чипсом
То же показано в исследованиях других видов. Сравнение 7 пар «городских» и «сельских» популяций больших синиц Parus major в разных районах Европы показывает, что среди первых преобладает стратегия разрешения стресса активным действием (в этом исследовании — связанного с поимкой). Удерживаемые в руках в течение 5 мин., они интенсивней вокализируют, больше вырываются и клюются. Напротив, вторые чаще «терпеливо переносят» поимку. Эти различия не зависят от местности или от возраста (Senar et al., 2017).
«Городские» особи везде отличаются от «сельских» или «лесных» конспецификов из региона не столько изменением «смелости», «интеллекта», других характеристик, составляющих поведенческий синдром (Sih et al., 2004; Sih, de Guidice, 2012), но
a) лучшим распознаванием ситуаций (разнообразие которых выше при городской жизни, как и качественные различия между ними), и
b) большей переключаемостью поведения от одной к другой, вопреки крайней «дробности» подразделения пространства-времени существования «городских» птиц на «участки», «захватываемые» каждой из них. Поэтому в «городских» популяциях разных видов исчезает скоррелированность форм поведения, из которых исследователь составляет синдром (высокозначимая вне города). Каждую из них урбанизация всё больше меняет направленно, хотя они всё более рассогласовываются друг относительно друга.
У части видов в начале урбанизации уровень кортикостерона неизменно растёт4, что скорее способствует её успеху (Bonier, 2012; Sepp et al., 2018, table 2). С другой стороны, сохранение повышенного уровня кортикостерона отнюдь не препятствует большей способности оценивания ситуации, прогнозирования изменений среды и другим когнитивным «достижениям» «городских» птиц, а скорей сопрягается с ним. У одомашненных видов, наоборот, то и другое связано отрицательно, и «достижения» возникают иначе.
Отбор на доброжелательность к людям, вольеру, вообще искусственному окружению блокирует запуск на них стресс-реакции, обычной для «диких» видов, снижает тревожность/агрессивность особей в ответ на новое, необычное в среде обитания, меняет отношение к нему на позитивное/добродушное сообразно уменьшению уровня глюкокортикоидов (Трут, 2007, 2008). Продлевается обучение молодых, что полней проявляет поведение, обычно подавленное восприятием всего связанного с людьми как опасности, в т.ч. исследование новых объектов, манипуляции с ними вместо бегства. Эти же изменения толкают использовать социальные «подсказки»: оба фактора увеличивают интеллект, оцениваемый решением проблемных задач (Hare et al., 2012; Хейр, Вудс, 2014).
У «городских» птиц отношение к новому и потенциально опасному вместо «доброжелательности» формируется «смелостью». Отнюдь не делаясь «добрей» и «доверчивей», они больше готовы к сближению с ним, и ситуативней меняют поведение сообразно контексту и последствиям подобной активности. Также растёт скорость сближения, особенно связанного с исследованием. Так, в «городских» популяциях больших синиц и черноголовых гаичек значимо преобладают «быстро обследующие» («fast» explorers). Носители этого типа индивидуальности, будучи помещёнными в новое пространство, осматривают его интенсивней (быстрей движутся, чаще перемещаются), но «поверхностней», чаще возвращаются к уже осмотренному.
Ему противопоставляются обследующие медленней, но намного тщательней («slow» explorers), преобладающие в исходных популяциях: первые смелей, агрессивней, но и стереотипней, склонны к копированию кормовых методов других птиц, вторые осторожны и робки, но инновативны; кормятся самостоятельно. Проблемные ситуации первые разрешают активным действием, вторые — пассивным переносом (проактивная vs реактивная тактика). См. Sih et al., 2004; Thompson et al., 2018).
Большая интенсивность перемещений, в т.ч. энергозатратных, вроде полётов и перепархиваний, увеличение порывистости каждого из них и зигзагообразности общей траектории прямо связано с лучшей ресурсообеспеченностью в городах (как минимум взрослых особей, у птенцов бывают проблемы с белковым кормом именно в нужный период из-за частых случаев меньшего обилия голых гусениц и другого массового корма в городских биотопах, Seress et al., 2018), резко расширившей возможность «активного бездельничанья». Эти черты индивидуальности наследственны — как и другие анализируемые в данной статье: отбор по ним в 4-х поколениях больших синиц был успешен и усилил их альтернативность Fidler et al., 2007; Riyahi et al., 2015, 2017; Baugh et al., 2017).
Скорей всего, в урбосреде важны оба варианта, но более выраженные, чем в исходных биотопах (отсюда отбор, производимый урбанизацией). У больших синиц и черноголовых гаичек постепенно вырисовывается связь альтернативы «быстрого» vs «медленного» исследования с альтернативой между скоростью и точностью решения проблемных задач и задач на социальное обучение (Lefebre, Aplin, 2017). У первых преобладает скорость, у вторых точность, почему внутри города они «сортируются» между разными элементами урболандшафта (Sprau, Dingemanse et al., 2017).
Наследуемость обоих вариантов индивидуальности также возрастает при урбанизации, в т.ч. потому что среди генов, влияющих на индивидуальность животного, наиболее важны кандидатные гены урбанизации SERT и DRD4, почему их считают «генами индивидуальности».

Волнистый астрильд
Последнее подтвердилось экспериментально. Использование антагонистов рецепторов дофамина для D1 и D2 метаболических путей (последний включает рецептор D4) показало немедленное изменение индивидуальности астрильдов Estrilda astrild после их кратковременного воздействия. Блокирование D2 уменьшает их социабельность (время, проводимое у зеркала в соответствующем тесте, и двигательную активность перед ним). Экспериментальное увеличение D2 действовало противоположно: повышало подвижность в социальном контексте (у зеркала), но снижало в прочих; D1 — увеличивало время, проводимое у кормушки, и интенсивность питания. Среди прочего эти манипуляции разрушили корреляции, исходно присутствующие у астрильдов: более подвижные птицы менее социабельны (Silva et al., 2020). Именно так разрушает урбанизация т. н. поведенческие синдромы, присутствующие у птиц в исходных популяциях: исчезновение связей между составляющими их качествами позволяет каждой из них при освоении данным видом данного города меняться независимо от всех прочих.
В силу всего вышесказанного — отнюдь не вследствие «более доброжелательного» отношения к людям и технике (т. е. синантропности, вовсе необязательной для урбанизации) — по сравнению с исходными популяциями у них сокращается средняя дистанция взлёта (Møller et al., 2012; Møller, 2014; Blumstein, 2014; Samia et al., 2017). Даниилом Т. Блюмштейном показано, что долговременное обитание по соседству с человеком (т. е. в условиях регулярного беспокойства), а отнюдь не конкретная опасность, следующая из встреч с людьми в этом районе (вроде охоты) увеличивает способность «диких» видов дифференцировать первое и второе, одновременно меняя реагирование на них. Беспокойство всё больше вызывает привыкание и ослабляет реакцию, вплоть до полного безразличия (англ. habituation), реальная опасность, напротив, более острую реакцию, то и другое меняется в целом пропорционально частоте и «тесноте» встреч с людьми.
Воспроизведение вокализаций хищников (вой шакалов) вместе с неопасными звуками (пение птиц) дикдикам Гюнтера Madoqua guentheri, воспринимающим опасность в первую очередь по звукам-предвестникам, показало зависимость их дифференциации от дистанции до человеческих поселений. Антилопы, обитавшие далее 500 м от них, равно настораживались и беспокоились при воспроизведении обоих звуков, жившие ближе их легко различали и реагировали дифференцировано. Сходным образом чернохвостый олень Odocoileus hemionus различает тревожные крики сурков и пение зонотрихий, «включает» оборонительное поведение в ответ на первые и остаётся безразличным ко вторым лишь при постоянном обитании рядом с людьми (Casco, Blumstein, 2012, цит.по: Blumstein, 2014).
Скажем, дистанция взлёта 8 околоводных видов птиц (солдатский чибис Vanellus miles, молуккский ибис Threskiornis molucca, белощёкая цапля Egretta novaehollandiae, крачка Берга Thalasseus bergii, пёстрый кулик-сорока Haematopus longirostris, австралийский пеликан Pelecanus conspicillatus, австралийская чайка Chroicocephalus novaehollandiae, малый веретенник Limosa lapponica) оцененная на 6 разных пляжах Ботанического залива близ Сиднея, в целом менялась обратно пропорционально их посещаемости людьми, т. е. общему беспокойству. Однако межвидовые различия в реактивности на него вполне сохранялись, и пересечения линий на графиках мало; дисперсионный анализ показывает, что отклик изменением дистанции взлёта зависит от вида5 и конкретного пляжа, но не взаимодействия этих факторов (Blumstein, 2014, fig.4.1).
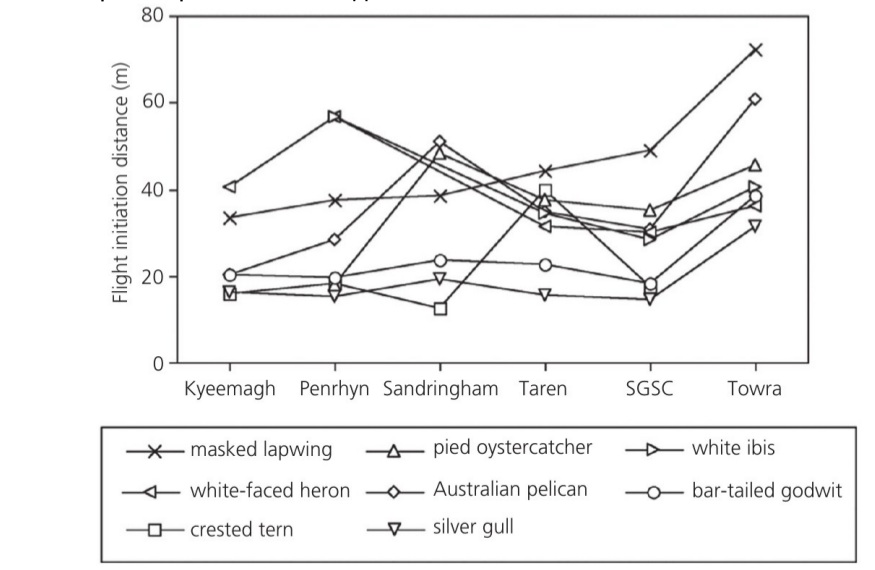
Рис.4.1. Ось Х — разные пляжи, ось У — дистанция взлёта
Посещаемость пляжа Санта-Моника в Калифорнии сильно разнилась по разные стороны от популярного причала: быстро падало в одну сторону по побережью и оставалось неизменно высоким в другую. Дистанция взлёта западных чаек L.occidentalis в первом случае была постоянной, во второй — увеличивалась пропорционально расстоянию от причала. Сходным образом действовал забор, деливший территорию водно-болотных угодий южной Калифорнии на участки, доступные экотуристам и нет. Дистанция взлёта во втором случае была выше (Blumstein, 2014, fig.4.3).
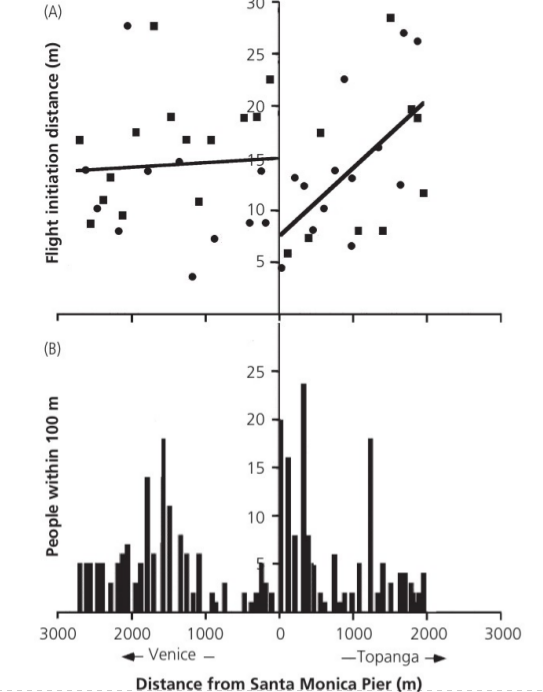
Рис.4.3. Дистанция взлёта западных чаек различно меняется в разные стороны от пляжа Санта-Моника
Лучшая дифференциация беспокойства от реальной опасности при урбанизации, кроме прогрессирующего привыкания, может вести и к противоположному — ещё большей чувствительности к нему, т. н. сенсибилизации. Это зависит от размера «островов» видовых биотопов, «архипелаг» которых создан их «охватом» и «раздроблением» растущим городским ареалом. В «точечных» местообитаниях, когда поселившимся птицам «некуда отступать», доминирует первое, на «островах», достаточно протяжённых для изоляции от беспокойства в «ядерной» части (или хуже доступных для посетителей, когда беспокойство осуществляется только с краёв, по дорожкам) — второе, хотя и здесь важны межвидовые различия (Blumstein, 2014).
Так, в окрестностях Лос-Анджелеса сохранившиеся участки водно-болотных угодий точечные, и обитающие там виды с ростом посещаемости демонстрируют привыкание. Рядом, в горах Санта-Моника 6 участков чаппараля достаточно протяжённы (разделены дорожками и пожарными проездами) и различались по посещаемости (2 с высокой и 4 — низкой). Из 14 исследованных здесь видов 10 не изменили дистанции взлёта с ростом посещаемости, а у 4 (американский чиж Spinus tristis, длиннохвостый крапивник Бьюика Thryomanes bewickii, темношапочная кустарниковая синица Psaltriparus minimus, крапивниковая синица Chamaea fasciata) она даже выросла (Blumstein, 2014, fig.4.4).
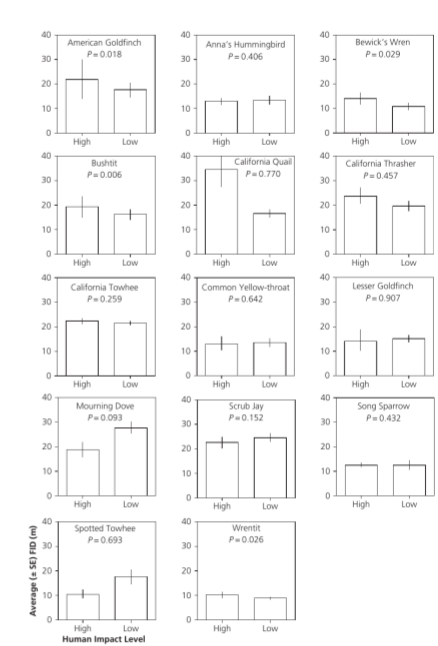
Рис.4.4
Возможность «отступить» от беспокойства толкает к появлению превентивно-защитных адаптаций (рост осторожности, заблаговременно исключающий контакт со стрессором). Поддерживая психологический комфорт птиц здесь и сейчас, они крайне вредны в долговременной перспективе (вид вытесняется из краевых зон каждого «острова», падает экологическая ёмкость местообитаний), особенно с учётом продолжающейся урбанизации и, соответственно, прогрессирующего расчленения, трансформации и сокращения «островов». И наоборот, точечность биотопов вызывает приспособительные изменения, рост безразличия к беспокойству, позволяющие существовать в ранее неблагоприятной среде, открывающие для вида новые территории, с ещё большим воздействием.
В разных странах Европы: Польше, Чехии, Италии, Франции исследовали дистанции взлёта в городских парках разной площади (n=79), с одной стороны, и на старых кладбищах (n=90), с другой, у 44 разных видов птиц. Все они на кладбищах подпускали наблюдателя ближе, чем в парках: это соотношение постоянно в разных странах. Дистанции взлёта (n=2139) везде были прямо пропорциональны размеру «зелёного острова» и обратно — плотности посетителей и/или могил (Morelli et al., 2018). Всё это подтверждает «принцип Ле Шателье» во взаимоотношениях «диких» видов с урболандшафтом, «наступающим» на их биотопы.
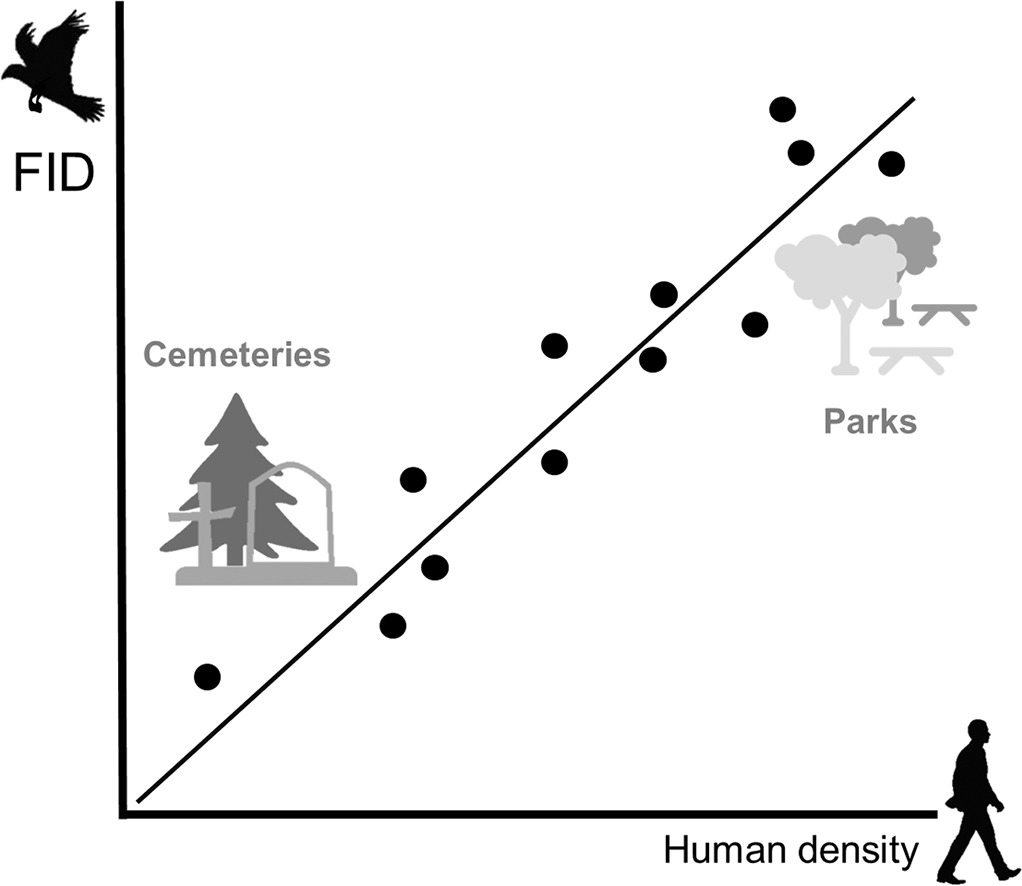 Для каждого вида наблюдаемая скорость антропогенной трансформации местообитаний и ландшафта в целом или:
Для каждого вида наблюдаемая скорость антропогенной трансформации местообитаний и ландшафта в целом или:
а) стимулирует его к трансформации популяционной системы в сторону более лабильных паттернов отношений, обеспечивающих устойчивость жизни в более фрагментированных, менее стабильных и меньших по площади «островах» местообитаний, чем было типично для вида ранее, или
б) стимулирует сокращение видовых популяций в районе соответствующих изменений с отступлением туда, где местообитания стабильней во времени и по конфигурации ближе к «материку». Иными словами, в ответ на преобразование природных ландшафтов растущей агломерацией, под действием пропорционального ему средового стресса разные виды птиц изменяются так, что оказываются нечувствительными к нему или уходят из места действия стрессора. Первое – возвратная урбанизация, второе – «отступление» на периферию, в обоих случаях проблема снимается (Фридман и др., 2016).
Добавим, что непосредственные проявления тревоги и бегства (дистанции взлёта, дистанция настораживания с фиксацией потенциальной опасности, частота осматриваний, прерывающих кормление или отдых, частота сердечных сокращений и пр.) в городских популяциях будут в среднем уменьшены по вполне объективной причине.
Прежде всего, в каждом из используемых биотопов урбосреда «предлагает» избыток ресурсов, «делая» бюджет времени и энергии менее напряжённым, оставляя больше того и другого на исследование окружения, отделение реальных опасностей от просто беспокоящих объектов, и выработку безразличия ко вторым. Избыток даёт возможность раньше обнаруживать и дольше прослеживаться первые, а вторые — подпускать ближе и быстрей вырабатывать безразличие, при лучшей дифференциации между категориями. Именно это мы и видим в действительности (Tätte et al., 2019; Gotanda et al., 2020).
Среди прочего, так «городские» птицы выигрывают ещё и от сокращения времязатрат на осматривания и ложные взлёты, существенного вследствие подъёма беспокойства при проникновении вида на сильнее застроенные районы города. Другой «дополнительный» выигрыш: этот избыток даёт больше времени для «активного бездельничанья». Оно же «скрытое», т. к. внешне напоминает агрессию, кормодобывание, спасение от опасности и т. д. виды активности, но при данной реализации носит нефункциональный характер, это игра, «сброс» избытка энергии и пр. Рост времязатрат на него разнообразит репертуар телодвижений и действий животного, способов манипуляции предметами, создаёт новые «инструменты» исследования окружения и «сырьё» для поведенческих инноваций (Дольник, 1986, 1995: 170-176; Мешкова, Фёдорович, 1996, приложение). Разнообразие моторных актов и видов деятельности птиц — важнейший предиктор инновативности и успеха решения проблемных задач, равно необходимых в освоении урбосреды (Lefebre, Aplin, 2017).
Напротив, «достижения» птиц из урбанизированных популяций — «переключение» между альтернативными формами поведения сообразно контексту, использование новизны в окружении (и, шире, других объектов и изменений в зоне досягаемости) в своих интересах, прогноз изменений урбосреды, — исключительно индивидуальны. Опираясь на них, они «следуют за средой» независимо друг от друга, особенно пребывая в «подвижном резерве» популяции (Фридман, Ерёмкин, 2009; Sol et al., 2013, 2016, 2020; Dukatez et al., 2017).
Ряд «городских» инноваций распространяется через социальное обучение (англ. cultural transmissiоn), от знаменитого прокалывания алюминиевых крышек молочных бутылок большими синицами и лазоревками6 до выучивания опасных людей (данные по американской вороне, Cornell et al., 2012 и многоголосым пересмешникам Mimus polyglottos, Levrey et al., 2009), решения проблемных задач вроде устранения преграды в доступе к корму или вариантов песни, меняющих частоту и амплитуду вокализации в сторону большей помехозащищённости. Индивиды, быстрей обучающиеся «копировать» чужое поведение, также быстрее решают проблемные задачи и оказываются инновативней (Lowry et al., 2013; Aplin et al., 2013, 2015; Sol et al., 2013, 2020; Lefebre, Aplin, 2017).
Однако выбор поведения в проблемных ситуациях, регулярно «создаваемых» городской жизнью, в каждом таком случае происходит строго индивидуально: более социальные виды не отличаются ни меньшей дистанцией взлёта в урболандшафтах, ни большей — в исходных биотопах, т. е. принять риск или спасаться, решает каждая птица в отдельности (Møller, 2009, 2014, сравни с: Фридман, 2016: 80-85). Здесь птицы могут подражать сородичам, другому виду, или использовать информацию о происходящем из их специфических сигналов, однако не побуждают их к совместному вниманию (англ. joint attention) к проблеме, либо совместному действию по её разрешению. Поэтому их нельзя гомологизовать с поиском «социальных подсказок» у доместицированных видов: собак, коз, лошадей и т. д., так же, как в случае с бонобо.
Развивающее воздействие урбосреды усиливается обстоятельствами, формирующими «плюсы» урбанизации для «диких» видов, задающими рост выживаемости в «городских» популяциях. Кроме новых видов местообитаний и кормовых методов, это более мягкий климат, временами отсутствие обычных хищников и специализированных конкурентов (Rebolo-Ifrán et al., 2017; Møller, Diaz, 2018; Фридман, 2018; Liker, 2020). Не везде — они урбанизируются не хуже жертв, как тетеревятник и поедаемые им вяхири, дрозды, пустельги, перепелятник (или серая неясыть Strix aluco) и певчие птицы. То же относится и к другим видам хищных птиц (от 1 до 10-15 видов в разных городах Европы, Северной Америки и Японии, Лыков, 2009, 2012; Фридман и др., 2016; Cooper et al., 2020) и млекопитающих (Santini et al., 2019).
В городе к ним добавляются кошки, собаки, разорители гнёзд вроде крыс, белок, серой вороны Corvus corone, сороки, сойки Garrulus glandarius (Луговой, 2002; Мацюра, Зимароева, 2016). Отсюда в зависимости от ситуации — конкретного города и региона с «его» спецификой урбанизации, способа размещения гнёзд, иногда вида птиц и пр. — риск гибели кладок или птенцов может расти или падать. См. данные по разорению искусственных гнёзд в г.Халапа — миллионнике, столице шт.Веракрус, Мексика (Rivera-López, MacGregor-Fors, 2017), естественным и искусственным гнёздам ( Vincsze et al., 2017) и др. Правда, в любом случае он не сильно меняется относительно такового в исходных местообитаниях, а большая часть отхода связана с размещением гнёзд в «экологических ловушках» (Фридман, Ерёмкин, 2009: 49 и разд.III.2): хищник лишь делает эту ошибку действительной.
В силу вышеописанного нельзя объяснять уменьшение дистанции взлёта «городских» птиц и другие изменения оборонительного поведения падением риска хищничества в городах, как часто делается (Møller, 2008, 2014; Møller et al., 2012). Изъятие обычного хищника из природной среды обитания разных видов, от гуппи Lebistes reticulatus до песочников Calidris bairdi и лосей Alces alces вызывает одни и те же микроэволюционные изменения, гомологичные росту доверчивости к человеку (бывшей обычной опасности) у беляевских лис, норок и пр. Там уменьшается настороженность, слабеет, потом исчезает реакция бегства и пр. Данные изменения необратимы; возвращение хищника после того, как они состоялись (реинтродукция волков Canis lupus и гризли Ursus arctos к лосям и оленям Йеллоустона) ведёт к катастрофической гибели животных. В природе (и с гуппи в эксперименте) оно часто заканчивалось вымиранием (Reznick et al., 2008).
Удаление хищника из среды обитания позволяет «сэкономить» на оборонительном поведении, и перестроить жизненный цикл: поднять среднеожидаемую продолжительность жизни (СОПЖ), уменьшить скороспелость особей и их вклад в каждое отдельное размножение, увеличить сложность и экспрессию форм социальной, в т.ч. брачной активности, также как затраты времени и энергии на неё (Reznick et al., 2008). То и другое присуще доместикации (Трут, 2007, 2008), хотя отбор, возникающий вследствие «маскировки» прежней опасности многажды возрастающим беспокойством, с одной стороны, и выработки дружелюбного отношения к ней, весьма различны. Первое отчасти сходно с перестройкой жизненного цикла в сторону «более К-» в городских популяциях птиц.
Поэтому изменения оборонительного поведения, отмеченные в городах, следствие отнюдь не уменьшения риска хищничества, но лучшей оценки ситуации, более точного переключения между альтернативными формами активности, выбора «запускаемого» поведения и лучшего «направления» его по цели (т. е. опять «развивающего» воздействия урбосреды). Так, метаанализ Samia et al. (2017) показывает уменьшение дистанции взлёта в «городских» популяциях сравнительно с исходными, и эти различия у 32 изученных видов увеличиваются на градиенте с юга на север Европы (3900 км, 92 попарных сравнений между популяциями). Чем северней, тем сильней сокращение дистанции взлёта при урбанизации. Однако пропорционально ему растут скорости бегства птиц от потенциальной опасности, и расстояние, на котором птица замечает её, настораживается и начинает отслеживать. Эта зависимость не отмечена у птиц из исходных популяций, притом что в обоих скорости бегства растут при движении в более высокие широты (Samia et al., 2017).
Как эти противоречивые сочетания реализованы в поведении, обсуждается ниже.
«Синдром одомашнивания»: бонобо выпадает из ряда
Все одомашненные виды (но не шимпанзе и бонобо) начинают решение с вовлечения «значимых других», рождающего «подсказки» (Хейр, Вудс, 2014: 46-62, 141-150). В отличие от диких предков, все они, попав в проблемную ситуацию, активно ищут «подсказок», и решают задачу на их основе совместно. Благодаря им собаки, козы, лошади и пр. в решении проблемных задач превосходят шимпанзе (Трут, 2007, 2008): это т. н. «кооперативность познания». См. связанные между собой различия домашних животных от их диких сородичей по морфологии, физиологии, поведению в Hare et al. (2012, table.1).
Лишь одна пара: «бонобо — обыкновенный шимпанзе» лишняя; увы, статья обосновывает как раз самодоместикацию P.paniscus — и ошибается. Авторы доказывают, что различиях между видами шимпанзе того же рода, что отличия домашней собаки от волка, домашней морской свинки — от дикой, лабораторных крыс и мышей — от пасюков и «диких» Mus musculus/M.domesticus, беляевских, доброжелательных к нам лис, — от диких родичей.
Что справедливо везде, кроме восприятия «социальных подсказок» со стороны людей и сородичей при решении когнитивных задач. Оба вида шимпанзе равно не обращают на них внимания, не ищут их и т.д., притом что существа весьма умные. Авторы утверждают обратное, однако работы, на которые они ссылаются (Herrmann et al., 2010; Hare, Kwetuenda, 2010; Wobber et al., 2010) совсем о другом: большей социальной связанности и социальной зависимости бонобо во всех контекстах взаимодействия. Если Pan troglodytes «бескорыстно» (вне контекста доминирования или возможности спаривания) делятся пищей с друзьями внутри «своей» группы, то бонобо — также с чужаками и с членами других групп (Fruth, Hohmann, 2018).
В решении же когнитивных задач оба вида равно нечувствительны к «подсказкам» и мало способны к совместному действию. А главное, что в понимании и использовании коммуникативных жестов бонобо не превосходит шимпанзе в той же степени, в какой собака — волка и другие пары в этой таблице (Herrmann et al., 2010, table 1; Hobaiter, Byrne, 2014; Graham et al., 2018). Новые данные по использованию орудий и хищничеству бонобо вообще ликвидировали различия между обоими видами (Fruth, Hohmann, 2018). Сегодня показано, насколько неверны утверждения, что P.paniscus лучше решает задачи, относящиеся к «теории сознания» и причинности в социальных взаимодействиях, тогда как P.troglodytes — к причинности физического мира и в инструментальной деятельности. У обоих видов миры социальных связей и физических явлений не отделены друг от друга, они равно способны строить планы и достигать результатов в том и другом, используя орудия или других особей, но «своими умом», без «подсказок» извне (Хейр, Вудс, 2014; Федорович, 2015).
Другой важный момент: бонобо архаичней Pan troglodytes по морфологии, особенностям онтогенеза, «конструкции» социальной системы. Первый ближе к общему предку рода Pan и «ветви», ведущей к австралопитекам и людям, второй — вид более продвинутый и молодой (Трут, 2008; Diogo et al., 2018; Fruth, Hohmann, 2018). Но во всех прочих парах в таблице доместицированный вид — потомок «дикого», не наоборот!
Наконец, размер мозга при одомашнивании уменьшается: он «может» миниатюризироваться благодаря «кооперативности познания» и роли «социальных подсказок» в решении проблемных задач (Hare et al., 2012, tabl.1; Дугаткин, Трут, 2019). Отсюда сомнения в применимости концепции «самоодомашнивания» к антропогенезу: в филогенетической линии, ведущей к Homo sapiens «напрямую», через гейдельбергского человека Homo heidelbergensis, без учёта «боковой ветви» к неандертальцу H.neanderthalensis c денисовцем H.denisovan, размер мозга, средний и относительный, только растёт, и лишь с раннего верхнего палеолита (27-25 тыс.лет назад) стал уменьшаться7. Однако существенный довод «за»: сравнение геномов человека разумного, неандертальца и денисовца показывает, что только у него шёл интенсивный отбор по генам, связанным с миграцией клеток нервного гребня, стержневого процесса в доместикации (Theofanopoulou et al., 2017). И к бонобо: его мозг не уменьшен сравнительно с шимпанзе.
Впрочем, есть отдельные сходства урбанизации и доместикации: продлённый период обучения, меняющий поведение так, как описано выше. Равно растут уровень базального метаболизма и эффективность усвоения пищи, интенсифицируется размножение. Виды с большим репродуктивным усилием (одно из отличий «-выскочек» и характеристик более лабильной стратегии) легче осваивают города сравнительно с близкими формами (Møller, Díaz, 2018), однако по мере успешного освоения их популяционные системы меняются в сторону «более -К». Однако эти сходства сугубо внешние, скрывающие за собой качественно различные изменения эволюционного характера — как и отдельные сходства в изменениях поведения при урбанизации, с одной стороны, и доместикации, с другой.
Список литературы
Багоцкая М. С., Смирнова А.А., Зорина З.А. Изучение способности серых ворон (Corvus cornix L.) решать задачи на добывание приманки из трубки с ловушкой// Журн. общ.биол. 2013. Т.74. №1. С.23-33.
Дольник В.Р. Изучение поведения животных с помощью анализа их бюджетов времени и энергии //Методы исследования в экологии и этологии. Пущино, 1986. С.94-106.
Дольник В.Р. Ресурсы времени и энергии у птиц в природе. (Труды ЗИН. т.179). СПб: Наука, 1995. 360 с.
Дугаткин Л., Трут Л.Н. Как приручить лису (и превратить её в собаку)? Сибирский эволюционный эксперимент. М.: Альпина Нон-фикшн, 2019. 290 с.
Луговой А.Е. Врановые Закарпатья в ХХ столетии // Экология врановых птиц в антропогенных ландшафтах. Мат-лы межд. конф. Саранск, 2002. C.83-86.
Лыков Е.Л. Фауна, население и экология гнездящихся птиц городов Центральной Европы (на примере Калининграда). Дисс.канд.биол.наук. М., 2009. 286 c.
Лыков Е.Л. Гнездящиеся дневные хищные птицы в городах Европы// Мат-лы VI Межд. конф. по соколообразным и совам Северной Евразии. Кривой Рог, 2012. С. 558-565.
Мацюра А.В., Зимароева А.А. Синантропизация врановых и особенности их адаптаций к антропогенным ландшафтам// Acta Biologica Sibirica. 2016. Т.2. №1. С.150-199.
Мешкова Н.Н., Федорович Е.Ю. Ориентировочно-исследовательская деятельность, подражание и игра как психологические механизмы адаптации высших позвоночных к урбанизированной среде. М.: Аргус, 1996. 226 c.
Обозова Т.А., Багоцкая М.С., Смирнова А.А., Зорина З.А. Сравнительная оценка способности разных видов птиц к решению протоорудийных задач// Зоол.ж. 2013. Т.92. №6. С.731-731.
Суслов В.В. Основное противоречие адаптации и пути его разрешения// Развитие жизни в процессе абиотических изменений на Земле. Иркутск: Байкальский музей Иркутского НЦ СО РАН (Листвянка), 2014. №3. С.102-103.
Трут Л.Н. Доместикация животных в историческом процессе и в эксперименте// Инф.вестн.ВОГИС. 2007. Т.11. №2. С.273-289.
Трут Л.Н. Эволюционные идеи Д.К. Беляева как концептуальный мост между биологией, социологией и медициной// Вавиловский журн. генетики и селекции. 2008. Т.12. №1-2. С.7-18.
Федорович Е.Ю. Социально-когнитивные способности антропоидов: эволюционные предпосылки нравственных основ сотрудничества у людей// Вопросы прикладной приматологии. Вып.2. Межвед. сб. науч. и науч.-метод. тр. М: Московский зоопарк, 2015. С.33-49.
Фридман В.С. Что не так с социобиологией?// Социально-экологические технологии. 2016. №4. С.78-115.
Фридман В.С. Город как заказник: (пока?) неиспользуемая возможность //Памяти Г.П. Гапочки/Под ред. Д.Д.Соколова. М.: Юрайт, 2018. С. 164-193.
Фридман В.С., Ерёмкин Г.С. Урбанизация «диких» видов птиц в контексте эволюции урболандшафта. М.: URSS, 2009. 240 c.
Фридман В.С., Ерёмкин Г.С., Захарова Н.Ю., Возвратная урбанизация – последний шанс на спасение уязвимых видов птиц Европы?// Russ. J. Ecosyst. Ecol. 2016. V.1. №4. http://rjee.ru/rjee-1-4-2016-3/
Хейр Б., Вудс В. Почему собаки гораздо умней, чем вы думаете. Спб: Питер, 2014. 145 c.
Aplin L.M., Sheldon B.C., Morand-Ferron J. Milk bottles revisited: social learning and individual variation in the blue tit, Cyanistes caeruleus// Anim.Behav. 2013. V.85. №6. Р.1225–1232.
Aplin L.M., Farine D.R., Morand-Ferron J. et al. Experimentally induced innovations lead to persistent culture via conformity in wild birds// Nature. 2015. V.518. №7540. Р.538–541.
Atwell J.W., Cardoso G.C., Whittaker D.J. et al. Boldness behavior and stress physiology in a novel urban environment suggest rapid correlated evolutionary adaptation// Behav.Ecol. 2012. V.23. №5. Р.960-969.
Batabyal A., Thaker M. Lizards from suburban areas learn faster to stay safe// Biol.lett. 2019. V.15. №2. P.20190009.
Baugh A.T., Witonsky K.R., Davidson S.C. et al. Novelty induces behavioural and glucocorticoid responses in a songbird artificially selected for divergent personalities// Anim.Beh. 2017. V.130. P.221-231.
Baxter-Gilbert J., Riley J.L., Whiting M.J. Bold New World: urbanization promotes an innate behavioral trait in a lizard// Behav.Ecol.Sociobiol. 2019. V.73. №8. P.105. https://doi.org/10.1007/s00265-019-2713-9
Blumstein D.T. Attention, habituation, and antipredator behaviour: implications for urban birds// Avian Urban Ecology. Behavioural and Physiological Adaptations. Eds. Diego Gil & Henrik Brumm. Oxford: Oxf.Univ.Press, 2014. P.41-53.
Bonier F. Hormones in the city: endocrine ecology of urban birds// Hormones and Behavior. 2012. V.61. №5. Р.763-772.
Carrete M., Tella J.L. Inter-individual variability in fear of humans and relative brain size of the species are related to contemporary urban invasion in birds// PLoS ONE. 2011. V.6. P.e18859. doi: 10.1371/journal.pone.0018859
Carrete M., Tella J.L. Behavioral correlations associated with fear of humans differ between rural and urban burrowing owls// Front.Ecol. Evolution. 2017. V.5. №54.https://doi.org/10.3389/fevo.2017.00054
Cornell H.N., Marzluff J.M., Pecoraro S. Social learning spreads knowledge about dangerous humans among American crows// Proc. Roy. Soc.y B. 2012. V.279. №1728. Р.499-508.
Davies S., Beck M.L., Sewall K.B. Territorial aggression in urban and rural Song Sparrows is correlated with corticosterone, but not testosterone// Horm. and beh. 2017. V.98. P.8-15.
Deviche P., Davies S. Reproductive phenology of urban birds: environmental cues and mechanisms// Avian Urban Ecology. Behavioural and Physiological Adaptations. Eds. Diego Gil & Henrik Brumm. Oxford: Oxf.Univ.Press, 2014. P.98-115.
Diaz M., Møller A.P., Flensted-Jensen E., et al. The geography of fear: a latitudinal gradient in anti-predator escape distances of birds across Europe// PLoS One 2013. V.8. №5. Р.e64634.
Diogo R., Molnar J.L., Wood B. Bonobo anatomy reveals stasis and mosaicism in chimpanzee evolution, and supports bonobos as the most appropriate extant mode l for the common ancestor of chimpanzees and humans// Sci.rep. 2017. V.7. №1. Р.1-8.
Ducatez S., Audet J.N., Rodriguez J.R. et al. Innovativeness and the effects of urbanization on risk-taking behaviors in wild Barbados birds// Anim.cogn. 2017. V.20. №1. Р.33-42.
Ducatez S., Sol D., Sayol F., Lefebvre L. Behavioural plasticity is associated with reduced extinction risk in birds // Nature Ecol.Evol. 2020. P.1-6. DOI: 10.1038/s41559-020-1168-8.
Esposito R.M.M.K. Being Neighborly: Costs and Benefits of Nesting Associations between American Crows (Corvus brachyrhynchos) and Black-Billed Magpies (Pica hudsonia). Thesis of PhD. Univ.Louis.at Lafayette, 2015. 209 pp.
Fidler A. E., van Oers K., Drent P.J. et al. Drd4 gene polymorphisms are associated with personality variation in a passerine bird// Proc.Roy.Soc. B. 2007. V.274. №1619. Р.1685-1691.
Fisher J. The collared turtle dove in Europe// British Birds. 1953. V.46. P.153-181.
Fisher J., Hinde R.A. The opening of milk bottles by birds // British Birds. 1949. V. 42. №11. P. 347–357
Fisher J., Hinde R.A. The Opening of Milk Bottles by Birds// Nature. 1952. V.169. P.1006-1007.
Fokidis H.B., Orchinik M., Deviche P. Context-specific territorial behavior in urban birds: No evidence for involvement of testosterone or corticosterone//Horm. and Beh. 2011. V.59. №1. Р.133–143.
Fruth B., Hohmann G. Food sharing across borders// Hum.Nat. 2018. V.29. №2. Р.91-103.
Gotanda K.M. Human influences on antipredator behaviour in Darwin’s finches// J.Anim. Ecol. 2020. V.89. №2. Р.614-622.
Graham K.E., Hobaiter C., Ounsley J. et.al. Bonobo and chimpanzee gestures overlap extensively in meaning// PLoS biology. 2018. V.16. №2. Р. E2004825.
Griffin A.S., Tebbich S., Bugnyar Th. Animal cognition in a human-dominated world// Anim. Cogn. 2017. V.20. P. 1-6.
Hare B., Kwetuenda S. Bonobos voluntarily share their own food with others// Curr. Biol. 2010. V.20. P.230e231.
Hare B., Wobber V., Wrangham R. The self-domestication hypothesis: evolution of bonobo psychology is due to selection against aggression// Anim.Behav. 2012. V.83. №3. Р.573-585.
Herrmann E., Hare B., Call J., Tomasello M. Differences in the cognitive skills of bonobos and chimpanzees// PloS One. 2010. V.5. P.e12438.
Hobaiter C., Byrne R.W. The meanings of chimpanzee gestures// Curr.Biol. 2014. V.24. №14. Р.1596-1600.
Kozlovsky D.Y., Weissgerber E.A., Pravosudov V.V. What makes specialized food-caching mountain chickadees successful city slickers?// Proc.Roy.Soc.B. 2017. V.284. №1855. Р.20162613.
Lapiedra O. Urban behavioral ecology: lessons from Anolis lizards// Integr.Comp. Biol. 2018. V.58. №5. Р.939-947.
Lapiedra O., Schoener T.W., Leal M. et al. Predator-driven natural selection on risk-taking behavior in anole lizards// Science. 2018. V.360. №6392. Р.1017-1020.
Lefebvre L., Aplin L.M. Social learning and innovation// Avian cognition. Chapt. 6. Cambridge: Cambridge Univ. Press, 2017. P.93-117.
Levey D.J., Londoño G.A., Ungvari-Martin J. et al. Urban mockingbirds quickly learn to identify individual humans// PNAS. 2009. V.106. №22. Р.8959-8962.
Liker A., Bókony V. Larger groups are more successful in innovative problem solving in house sparrows// PNAS. 2009. V.106. №19. Р.7893-7898.
Liker A. Biologia Futura: adaptive changes in urban populations// Biologia Futura. 2020. V.71. P.1-8.
Lowry H., Lill A., Wong B.B. Behavioural responses of wildlife to urban environments// Biol.rev. 2013. V.88. №3. Р.537-549.
De Meester G., Lambreghts Y., Briesen B. et al. Hunt or hide: How insularity and urbanization affect foraging decisions in lizards// Ethology. 2018. V.124. №4. Р.227-235.
Miranda A.C., Schielzeth H., Sonntag T., Partecke J. Urbanization and its effects on personality traits: a result of microevolution or phenotypic plasticity?// Glob. change biol. 2013. V.19. №9. Р.2634-2644.
Miranda, A.C. Mechanisms of Behavioural Change in Urban Animals: The Role of Microevolution and Phenotypic Plasticity// Ecol.Conserv. Birds in Urb.Envir. Eds. E.Murgui, M.Hedblom. Springer Int. Publ., 2017. Р.113–132. doi:10.1007/978-3-319-43314-1_7
Møller A.P. Flight distance of urban birds, predation and selection for urban life// Behav. Ecol.Sociobiol. 2008. V.63. P.63 – 75.
Møller A.P. Successful city dwellers: A comparative study of the ecological characteristics of urban birds in the Western Palearctic// Oecologia. 2009. V.159. P.849 – 858.
Møller A.P. Interspecific variation in fear responses predicts urbanization in birds// Behav. Ecol. 2010. V.21. №2. Р.365-371.
Møller A.P. Behavioural and ecological predictors of urbanization//Avian Urban Ecology. Behavioural and Physiological Adaptations. Eds. Diego Gil & Henrik Brumm. Oxford: Oxf.Univ.Press, 2014. P.54-68.
Møller AP, Ibáñez-Álamo J.D. Escape behavior of birds provides evidence of predation being involved in urbanization// Anim. Behav. 2012. V.84. P.341–348.
Møller, A.P., Diaz M., Flensted-Jensen E. et al. High urban population density of birds reflects their timing of urbanization// Oecologia. 2012. V.170. №3. Р.867-875
Morelli F., Mikula P., Benedetti Y. et al. Escape behaviour of birds in urban parks and cemeteries across Europe: Evidence of behavioural adaptation to human activity// Sci. total env. 2018. V.631. P.803-810.
Partecke J. Mechanisms of phenotypic responses following colonization of urban areas: from plastic to genetic adaptation// Avian Urban Ecology. Behavioural and Physiological Adaptations. Eds. Diego Gil & Henrik Brumm. Oxford: Oxf.Univ.Press, 2014. P.131-142.
Pellitteri-Rosa D., Bellati A., Cocca W. et al. Urbanization affects refuge use and habituation to predators in a polymorphic lizard// Anim. Behav. 2017. V.123. P.359-367.
Preiszner B., Papp S., Pipoly I. et al. Problem-solving performance and reproductive success of great tits in urban and forest habitats// Anim.cogn. 2017. V.20. №1. Р.53-63.
Rebolo-Ifrán N., Carrete M., Sanz-Aguilar A. et al. Links between fear of humans, stress and survival support a non-random distribution of birds among urban and rural habitats// Sci.Rep. 2015. V.5. P.13723.
Rebolo-Ifrán N., Tella J.L., Carrete M. Urban conservation hotspots: predation release allows the grassland-specialist burrowing owl to perform better in the city// Scient.Rep. 2017. V.7. №1. Р.1-9.
Reznick D. N., Ghalambor C.K., Crooks K. Understanding the Evolutionary Consequences of Predator Removal in Natural Communities: Experimental Studies of Evolution in Guppies as a Model System// Mol.Ecol. 2008. V.17. P.97-107.
Rivera-López A., MacGregor-Fors I. Urban predation: a case study assessing artificial nest survival in a neotropical city// Urban Ecosystems. 2016. V.19. №2. Р.649–655.
Riyahi S., Sánchez-Delgado M., Calafell F. et al. Combined epigenetic and intraspecific variation of the DRD4 and SERT genes influence novelty seeking behavior in great tit Parus major// Epigenetics. 2015. V.10. №6. Р.516-525.
Samia D. S., Blumstein D.T., Díaz M. et al. Rural-urban differences in escape behavior of European birds across a latitudinal gradient// Front.Ecol.Evol. 2017. V.5. P.66. https://doi.org/10.3389/fevo.2017.00066
Santini L., González‐Suárez M., Russo D. et al. One strategy does not fit all: determinants of urban adaptation in mammals// Ecol.lett. 2019. V.22. №2. Р.365-376.
Senar J.C., Garamszegi L.Z., Tilgar V. et al. Urban great tits (Parus major) show higher distress calling and pecking rates than rural birds across Europe// Front. Ecol. Evol. 2017. V.5. P.163. https://doi.org/10.3389/fevo.2017.00163
Sepp T., McGraw K.J., Kaasik A. et al. A review of urban impacts on avian life‐history evolution: Does city living lead to slower pace of life?// Glob. change biol. 2018. V.24. №4. Р. 1452-1469.
Sepp T., McGraw K. J., Giraudeau M. Urban Sexual Selection// Urban Evolutionary Biology. M. Szulkin, J. Munshi-South, A. Charmantier (Ed.). Oxf.: Oxf. Univ.y Press, 2020. P.234-250
Seress G., Hammer T., Bókony V. et al. Impact of urbanization on abundance and phenology of caterpillars and consequences for breeding in an insectivorous bird// Ecol. Appl. 2018. V.28. P.5. S.1143-1156.
Sih A., Bell A., Johnson J.C. Behavioral syndromes: an ecological and evolutionary overview// Trend. ecol. evol. 2004. V.19. №7. Р.372-378.
Sih A., del Giudice M. Linking behavioural syndromes and cognition: a behavioural ecology perspective// Phil.Trans. R. Soc.B. 2012. V.367. P.2762–2772.
Silva P.A., Trigo S., Marques C.I. et al. Experimental evidence for a role of dopamine in avian personality traits// J. Exp. Biol. 2020. V.223. №3. https://jeb.biologists.org/content/223/3/jeb216499
Sol D., Lapiedra O., González-Lagos C. Behavioural adjustments for a life in the city// Anim. Behav. 2013. V.85. №5. Р.1101-1112.
Sol D., Sayol F., Ducatez S., Lefebvre L. The life-history basis of behavioural innovations// Phil.Transact.Roy.Soc. B. 2016. V.371. №1690. Р.20150187
Sol D., Lapiedra O., Ducatez S. Cognition and Adaptation to Urban// Urban Evolutionary Biology. M. Szulkin, J. Munshi-South, A. Charmantier (Ed.). Oxf.: Oxf. Univ.y Press, 2020. Chapt.15. P.253-265.
Sprau P., Dingemanse N.J. An approach to distinguish between plasticity and non-random distributions of behavioral types along urban gradients in a wild passerine bird// Front.Ecol.Evol. 2017. V.5. №92.
Suslov V.V. Par force evolution as a mechanism for rapid adaptation// Paleontological Journal. 2013. V.47. №9. Р.1048-1055.
Tätte K., Ibáñez-Álamo J.D., Markó G. et al. Antipredator function of vigilance re-examined: vigilant birds delay escape// Anim.Behav. 2019. V.156. P.97-110.
Theofanopoulou C., Gastaldon S., O’Rourke T., et al. Self-domestication in Homo sapiens: Insights from comparative genomics// PloS Оne. 2017. V.12. №10. Р.e0185306.
Thompson M.J., Evans J.C., Parsons S. et al. Urbanization and individual differences in exploration and plasticity// Behav.Ecol. 2018. V.29. №6. Р.1415-1425.
Vincze E., Seress G., Lagisz M. et al. Does urbanization affect predation of bird nests? A meta-analysis// Front.Ecol.Evol. 2017. V.5. P.29. https://doi.org/10.3389/fevo.2017.00029
Walsh S., Goulet C.T., Wong B.B. et al. Inherent behavioural traits enable a widespread lizard to cope with urban life// J.Zool. 2018. V.306. №3. Р.189-196.
Wobber V., Wrangham R., Hare B. Bonobos exhibit delayed development of social behavior and cognition relative to chimpanzees// Curr.Biol. 2010b. V.20. P.226e230
Примечания
Свежие комментарии