 Print
Print  PDF
PDF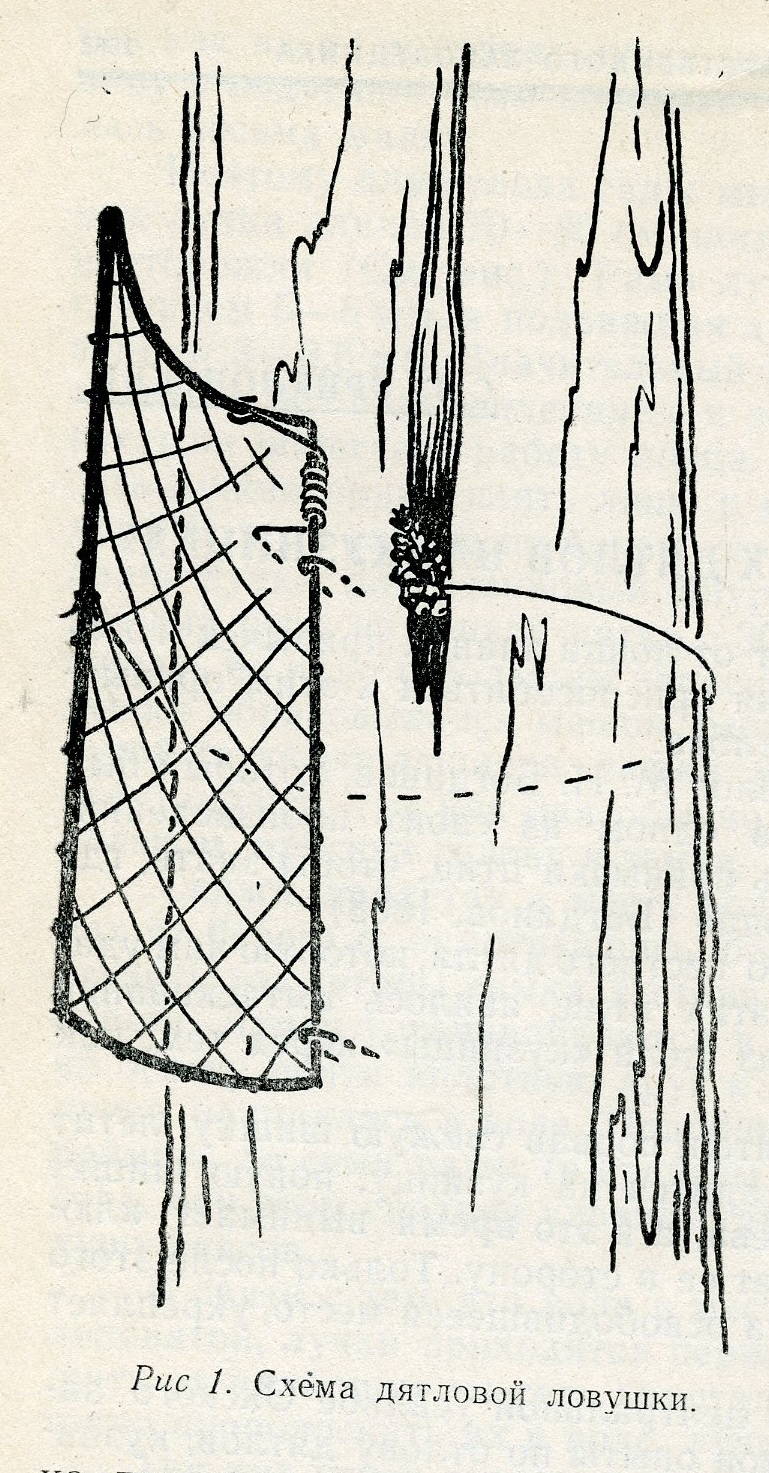
Введение
Содержание
Красивый пример инстинктивного действия (и чем оно отличается от гибкой, пластичной повседневной активности) мы видим в осенне-зимнем долблении большим пёстрым дятлом шишек сосны и ели на «кузницах». Чем управляется эта активность? Ключевым раздражителем в виде старой шишки в «станке» из которой птицей уже выбраны все семена (поскольку неотогнутые чешуи по мере долбления попадаются всё реже и реже, и шишку приходится всё чаще вертеть в «станке», чтобы попасть на неподолбленный бок).
Когда это произошло, дятел не может не удержаться, чтоб не слететь за новой, а, принеся её, обязательно прижмёт новую грудью и дёрнет клювом за старую шишку, чтобы вынуть.
Это инстинкт, автоматичный настолько, что используется для отлова дятлов на «кузницах«. Орнитологи настораживают лучок за старую шишку а дятел, прилетев, дергает за неё и спускает сторожок (Приклонский, 1962). Он не может не дёрнуть, хотя видит это странное сооружение; дёргает даже в случае, когда уже попадался и пр. См. статью С.Г.Приклонского ниже.
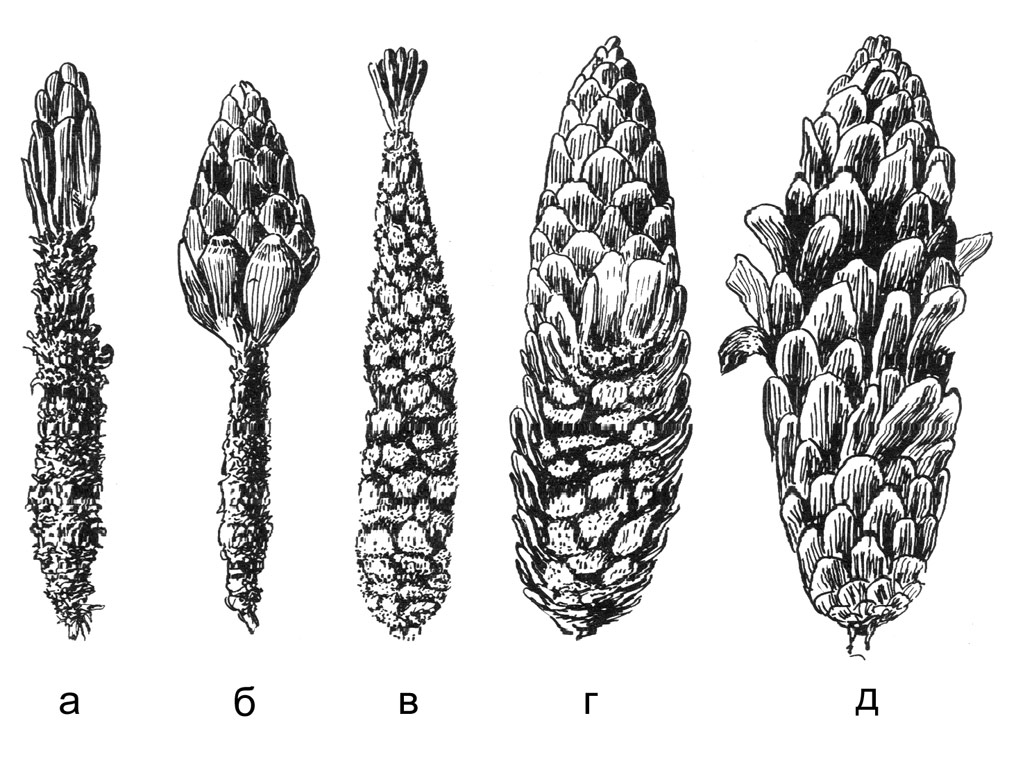
Еловые шишки, из которых извлекались семена разными зверями и птицами: а, б — белкой; в, г — лесными мышами; д — большим пестрым дятлом.
Отлов больших пёстрых дятлов на «кузницах»
Успешная ловля птиц требует от ловца знания привычек и особенностей поведения птиц, умения приспособиться к этим привычкам и использовать их в целях отлова.
Выдающийся русский орнитолог М. Н. Богданов, который был прекрасным птицеловом, писал в одном из своих произведений: «Главное дело в том, чтобы знать привычки птиц, чтобы знать, где какую ловить и в какое время года» (Богданов, 1888).
Такой «привычкой» большого пестрого дятла, которую нам удалось использовать для отлова этих.птиц, явилось вытаскивание старой «обработанной» шишки из щели «кузницы» перед тем, как он вставляет на ее место новую.
Обычно большой пестрый дятел, сорвав свежую шишку, летит держа ее в клюве, к кузнице; садится на кузницу, новую шишку прижимает грудью к стволу дерева, а в это время вынимает клювом старую шишку и отбрасывает ее в сторону. Только после этого дятел вставляет новую шишку на освободившееся место, укрепляет ее и начинает расклевывать.
В участке соснового бора в центральной усадьбе Окского заповедника, где мы проводили свои опыты по отлову дятлов, кузницы в большинстве случаев расположены в сухих вершинах на высоте 5—18 м от земли. Кузницы представляют собой выдолбленную в сухой древесине щель длиной 12—35 см, шириной 2—3,5 см. Глубина щели достигает в ряде случаев 5 см, но чаще всего бывает равна 2,5—3 см. Иногда дятлы делают кузницу, зажимая шишку между сучками или в расщелину, образовавшуюся после слома вершины дерева. Но такие кузницы редки. В обследованном нами участке леса площадью в 4 га из 36 обнаруженных там кузниц 30 были выдолблены дятлом в стволах деревьев и лишь 6, то есть всего около 17%, оказались «естественными». Обычно кузницу легко обнаружить по большому количеству раздолбленных шишек, валяющихся под ней на земле.
 Мы отлавливали дятлов только на выдолбленных ими кузницах, на стволах деревьев не тоньше 12 см. Наша ловушка представляет собой прямоугольную проволочную рамку размером 28X13 см, затянутую сеткой с ячеей 20—25 мм, имеющую пружину, один конец которой закреплен на горизонтальной стороне рамки, а второй свободен. Осью пружины служит вертикальная сторона рамки (рис. 1). Проволока для рамки может быть любая — сечением от 2,5 до 4 мм; для пружины мы брали сталистую проволоку сечением 1,6 мм. К свободной от пружины вертикальной стороне рамки привязывается прочная нитка длиной 70—80 см.
Мы отлавливали дятлов только на выдолбленных ими кузницах, на стволах деревьев не тоньше 12 см. Наша ловушка представляет собой прямоугольную проволочную рамку размером 28X13 см, затянутую сеткой с ячеей 20—25 мм, имеющую пружину, один конец которой закреплен на горизонтальной стороне рамки, а второй свободен. Осью пружины служит вертикальная сторона рамки (рис. 1). Проволока для рамки может быть любая — сечением от 2,5 до 4 мм; для пружины мы брали сталистую проволоку сечением 1,6 мм. К свободной от пружины вертикальной стороне рамки привязывается прочная нитка длиной 70—80 см.
Установка ловушки производится следующим образом. Рамка примеряется к кузнице, и ее горизонтальные стороны изгибаются таким образом, чтобы рамка плотно облегала дерево. Затем вертикальная сторона, служившая осью пружины, прикрепляется к дереву двумя проволочными шпильками длиной 2,5—3 см, которые забиваются молотком на расстоянии 3—4 см от .ближайшего края кузницы (перед этим свободный конец пружины заводится в сторону, противоположную кузнице).
Затем нитку обводят вокруг дерева позади кузницы и, натягивая, открывают ловушку, а в том месте, где нитка доходит до дальнего от осевой стороны рамки края кузницы, завязывают узел или привязывают небольшой кусочек дерева, который зажимается раздолбленной уже шишкой, находившейся в кузнице перед установкой ловушки.
Большие пестрые дятлы обычно наиболее активно «работают» на кузницах с утра до 12—13 часов, поэтому замечать кузницы, где дятлы бывают чаще, следует утром. Устанавливать же ловушки лучше во второй половине дня или вечером, выбирая время, когда дятла нет поблизости. В тех случаях, когда дятлы присутствовали при установке ловушки, чаще всего ловушки пустовали по нескольку дней. Очевидно, дятлы пугались человека, устанавливавшего ловушки, и улетали на другую кузницу.
Отлов дятлов мы проводили с 12 по 19 ноября 1957 г. и в конце января 1958 г. на 19 кузницах. В период отлова на этом участке постоянно держалось от 5 до 8 дятлов. С помощью окраски дятлов установлено, что каждый из них расклевывал шишки на 2—5 кузницах, находившихся на участке, площадь которого в среднем не превышала одного та. Одновременно выставлялось до 5 ловушек. Уловистость ловушек в наших опытах не превышала 5 птиц на 100 ловушко-часов.

Всего на 10 кузницах было поймано 13 дятлов (из них 9 самцов) и один поползень. Интересно отметить, что на кузнице № 5 было отловлено три, а на кузнице № 14 — два дятла. При этом два первых дятла ловились в один и тот же день через 4—6 часов один после другого. Все дятлы, пойманные на этих кузницах, — самцы. На кузнице № 1 12.XI.- 1957 г. был пойман дятел с кольцом F—178690, помеченный на усадьбе заповедника 22.XII.-1953 г. и ловившийся повторно 17.1.-1954 г. (рис. 2).
Таким образом, уже первые опыты по отлову дятлов на кузницах дали интересные результаты. Выяснено, что дятлы «работают» на нескольких кузницах, в то же время одна кузница может принадлежать нескольким дятлам. С помощью отлова дятлов на кузницах можно увеличить процент возврата колец в местах массового кольцевания этих птиц, ибо других способов отлова дятлов пока не существует. (Взрослых дятлов отлавливали случайно на гнездах, при ночных осмотрах скворечников (Сапетин, Карпович, 1956). Случайно они попадаются и в другие ловушки.
Но самым важным выводом, несомненно, является то, что для успешной ловли птиц нужно хорошее знание их поведения. «Это знание дается только тем, кто сам следит за птицами, внимательно изучает их привычки, их образ жизни» (Богданов, 1888).
ЛИТЕРАТУРА
1. Богданов М. Н., 1888. Из жизни русской природы.
2. Карпович В. Н. и Сапетин Я. В., 1956. О роли искусственных гнездовий для птиц в зимнее время. Сб. Пути и методы использования птиц для борьбы с вредными насекомыми.
Труды Окского зап-ка. Т.4. Вологда, 1962. С.427-430
Сходный опыт был поставлен на золотом дятле Colaptes auratus Ноблом в 1936 г. (рис.4, 5, 7, 8 — из этой работы). Самец C.auratus отличается от самки наличием чёрных «усов».
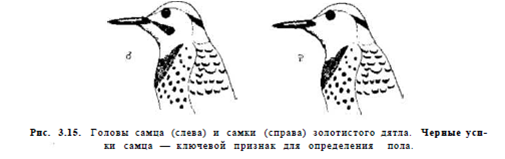
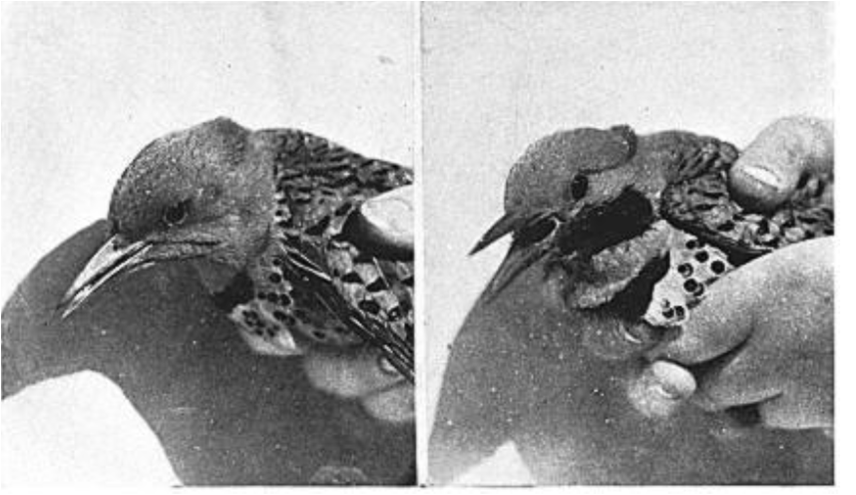
Рис.4
После того как образовалась пара и вроде бы отношения партнёров установились, Нобл отловил самку и приклеил «усики» (рис.4). Самец напал на неё и выгнал (см. рис.5сверху, угрожающий самец слева, исполняет типичные демонстрации ритуализованной угрозы самцов; «перекрашенная» самка справа, возбуждена и стрессирована неожиданным нападением, что показывает раскрытый и повёрнутый на сторону хвост). После отклеивания «усов» самец немедленно принял самку обратно, и ухаживал за ней, и не угрожал (см.рис.5снизу).

Рис.5
Но! Самец более чем «осознавал», что перед ним — его прежняя подруга, и тем не менее «не мог удержаться».
Как это доказать? Тут нужен небольшой экскурс в организацию социальной жизни золотых дятлов. Самцы прилетают раньше и делят территорию, их поведение в конфликтах друг с другом высокоритуализовано — см. взаимодействия на границах, где демонстрируются именно эти самые «усы», также как у большого пёстрого, белоспинного и других пёстрых дятлов (рис.6).

рис.6. Источник
Самки, прилетающие несколько позже, конкурируют не за территории, а за распределивших её самцов. Как у наших больших пёстрых дятлов, у золотых они доминируют в паре (причём в значительно большей степени — настолько, что можно говорить о реверсии половых ролей, когда на самца сваливается большая часть «работы» сперва по охране территории пары, потом по насиживанию и выкармливанию потомства, тем более что и полиандрия встречается часто, 4 группы на 71 моногамных).
В период ухаживания самки активно сближаются с самцами, и, столкнувшись в этом процессе друг с другом, активно дерутся (рис.7; самец справа равнодушно ждёт исхода конфликта, после чего ухаживает за победительницей, рис.7, самец слева, весь вытянувшись и плоско раскрыв хвост).

Но! Драки самок в отличие от самцов, не ритуализованы, наблюдаемые экспрессивные действия — раскрытый и повёрнутый в сторону хвост, семантически пусты, т.е. не «выражают» угрозы, но лишь общее возбуждение (см. рис. драк самок). Так вот, самец, угрожая «переделанной» самке как самцу, выказывает явный стресс с возбуждением (см. ещё раз рис.5а), чего не видим в случаях с настоящими самцами. Так сказать, мучается, но подчиняется действию релизера, как и должно быть в случае видового инстинкта, независимого от разумения и опыта, каковые индивидуальны.
Ещё одно важное замечание. Опыт Нобла удался лишь потому, что у золотого дятла формальная, а не настоящая моногамия (о них см. «Против «обмена еды на секс»). Самец и самка просто заняли одно территорию в результате распределения в пространстве, организуемого разными конкурентными процессами, у каждого пола в отдельности (рис.8; слева — самец угрожает приземлившемуся самцу, справа — самка возбуждённо атакует другую самку). Поэтому они (то бишь процессы) у самцов и самок управляются разными демонстрациями и при охране территории пары от нарушителей самка строго атакует самок, самец — самцов, о чём пишут и Нобл, и исследовавший их позже Килхэм.

Рис.8
Устойчивых брачных связей между партнёрами, как у сорок или прерийных полёвок, здесь нет (нет и взаимодействий, поддерживающих единство пары, вроде дуэтов и аллопрининга), хотя в 1960е годы Шортом показано, что партнёры отлично распознают друг друга по тонким вариациям голоса, тем более что призывом к спариванию служит специфический крик самки, принявшей позу подставления. Поэтому самец при изменении облика самки без проблем выгоняет её, потом принимает, в известном смысле она для него — не индивид, а комплекс ощущений.
При настоящей моногамии устойчивость брачной связи партнёров обеспечивает резистентность обоих к токованию «третьих» особей (как минимум пока он или она находятся рядом с тем, кто бы мог «соблазниться»). Изменение ключевых характеристик облика самца или самки здесь или ничего не меняет, пара сохраняется, или выгнанного партнёра уже не принять, оставшийся ведёт себя как холостой и ищет новую.
«Ум» и инстинкт
Ещё Л.В.Крушинским показано, что дятлы — умные птицы, с высокоразвитой рассудочной деятельностью (что подтверждается сейчас и нейроанатомией).
«Если щель в стволе дерева не подходит для закрепления в ней шишки, дятел соответствующим образом обрабатывает ее и подгоняет по размеру, причем для сосновых шишек он делает станок одной формы, а для еловых — другой. В одном дереве бывает два станка для разных шишек. Станки пробовали забивать фанерой. В этой новой поверхности дятел продалбливает точно такое же отверстие, как первоначальное. Описано похожее поведение дятлов при раздалбливании летков в скворечниках, которые они используют для ночлега. Дятел точно подгоняет размеры летка по своему росту, несколько раз делая «примерку». Вся эта сложная и разнообразная деятельность, несомненно, оставляет место и для проявления элементарного рассудка».
Л.В.Крушинский, 1986. Биологические основы рассудочной деятельности. 2е изд. М.: изд-во МГУ, 1986
Однако инстинкт и «ум», даже самый развитый, оказываются «параллельны». Как бы ни был развит второй, он, говоря языком феодальных отношений в Московской Руси, «не вступается», в область действия первого. Даже у таких умных тварей как мартышки Диана, инстинкт неизменно «слеп», и не может быть обогащён, а тем более изменён, результатами рассудочной деятельности и прошлого опыта. И наоборот: инстинкт облегчает обучение действиям «в том же направлении»или использующим инстинктивные акты (т.н. «инстинктивное смещение» Брелендов), но не добавляет им точности с автоматичностью.
Другой важный аспект «параллельности»: видовой инстинкт и индивидуальный «ум» (рассудочная деятельность, опыт, разные формы обучения) могут решать одни и те же задачи, со сравнимой эффективностью, но по-разному.
Уже лет 40 назад была показана способность птиц к «автоматическому» распознаванию образов (геометрических форм), при отсутствии подобных автоматизмов у человека, причём показана на одной и той же экспериментальной установке. Думаю, то же было бы при тестировании других низших позвоночных – и очень жаль что попробовали только с голубями.
Голубей в камере Скиннера обучали различать прямое и зеркальное изображение фигур, которые предъявлялись птицам ориентированными различным образом. Измеряли время реакции голубя в теста на инвариантность фигуры по отношению к повороту. Птицы были обучены указывать, какая из двух предъявленных для сравнения фигур более похожа на образец; в разных сериях опытов поворачивали на 00, 450 и 1800.
Когда закончились эксперименты с голубями, камеру демонтировали, и ту же самую тестовую панель с соответствующими зрительными сигналами и клавишами использовали в аналогичном тестировании людей.

Экспериментальная установка (вверху), с помощью которой голубям демонстрировали различные символические изображения (внизу). Птицы были обучены указывать, какая из двух предъявляемых для сравнения фигур больше всего похода на образец. Предъявляемые для сравнения фигуры были повернуты соответственно на 0, 45 и 180°. (Hollard, Delius, 1983.)
Соответственно, можно было использовать одни и те же комбинации стимулов для оценки способа решения данной задачи голубями и людьми. Процент ошибок при решении таких задач распознавания образов у людей и птиц был примерно одинаковым. Однако у людей необходимое время выбора верного ответа с ростом углового несоответствия между сопоставляемыми фигурами увеличивается прямо пропорционально последнему (Cooper, 1982), но у голубей время реакции от угла поворота фигуры никак не зависит (Hollard, Delius, 1983).
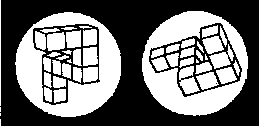
Пример пары фигур, которые используются для «психологической хронометрии». Это идентичные фигуры, но как бы повернутые на 80° одна относительно другой. (Shepard, Metzler, 1971.)
В обоих опытах верная реакция связана с тем, что реальная фигура сопоставляется с образом, существующим в «представлении» индивида. Кавычки поставлены потому, что результаты опыта скорее отрицают, чем подтверждают тот факт, что обработка визуальной информации происходит через формирование психических представлений, хотя некая модель для сопоставления должна быть обязательно.
То есть у голубей нужная реакция происходит помимо индивидуальных усилий животного, как бы «встроенным» в него механизмом видового уровня, возможно даже без образования психических представлений о распознаваемых фигурах (которые могло бы использовать животное, корректируя непосредственный опыт при решении подобных задач). Именно этот «автоматизм реагирования» в ответ на некие видоспецифические стимулы, заданные «идеальным образцом» имел в виду Лоренц, когда создавал понятие инстинкта и врождённого разрешающего механизмов, реализующего инстинкт во взаимодействии животных друг с другом (Lorenz, 1937а, b).
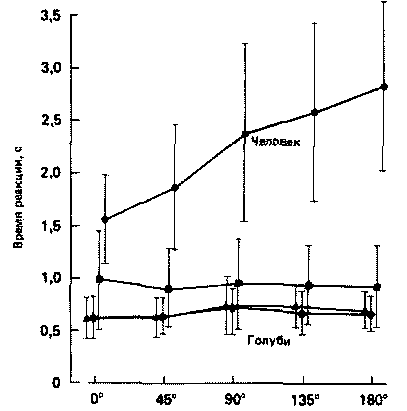
Среднее время реакции испытуемого как функция угла поворота предъявляемых для сравнения фигур (по результатам исследования 9 голубей и 22 человек). Обратите внимание на то, что у человека время реакции возрастает при увеличении угла поворота предъявляемых фигур, а у голубей — нет. (Hollard, Delius, 1983.)
У людей, в противоположность голубям, отсутствует эта инстинктивность реагирования на внешний стимул, по крайней мере, в данном опыте. В отличие от голубей люди решают задачу, создавая собственные психические образы (своего рода «общие идеи»), объекта, с которыми и происходит сопоставление: задержка связана с мысленным вращением сопоставляемого образа, которое требует времени (примерно 30 мс на каждые 200, Cooper, 1982).
Голуби используют врождённые схемы реагирования как механизм для распознавания и отреагирования объектов, значимость которых определяется неинстинктивно, процессами обучения. Именно так Конрад Лоренц понимал роль инстинктов в своей «естественной истории познания», в филогенетическом развитии способности к познанию мира. Инстинкты суть видоспецифические механизмы, «наводящие» индивидуальное познание (обучение, рассудочную деятельность, интеллект, другие когнитивные способности) именно на те элементы внешнего мира, которые значимы с точки образа жизни данного вида, свойственной практики взаимодействия индивидов друг с другом и с внешней средой в борьбе за существование (Лоренц, 1998).
У людей (и человекообразных обезьян) индивидуальная способность к познанию мира достигает настолько высокой степени развития (вместе с механизмами индивидуальной трансляции, способными «тиражировать» познанное с воспроизведением нужных решений в нужной ситуации по «идеальному образцу»), что система наведения, связанная с инстинктами, оказывается ненужной и даже вредной.
Данное заключение, при всей его «еретичности» — единственный способ объяснить противоположные результаты опытов с голубями и людьми на основе одной и той же концепции (что очень важно).
То есть человек решает задачу мысленным сопоставлением фигур, «вращением» их в уме с созданием соответствующего психического образа, принимает решение и делает выводы, поэтому время задержки пропорционально трудности задачи. Голубь же ничего не решает, его психика (которая несомненно есть, как и интеллект, и способность к обучению) никак не задействована в решении этой задачи, индивид здесь – чистая (ре)активность, которая предоставляет себя «в распоряжение» врождённым схемам реагирования вида (а не индивида, индивид ими только пользуется и платит за это неким умалением возможностей собственной психики, относящейся именно к индивидуальному уровню).
Понятно, что этологическому определению инстинкта соответствует именно и только второй случай, и всякое «расширение» понятия инстинкта достигается за счёт смешения «инстинкта» (как врождённого механизма реагирования) и «стереотипа» (как всяких ригидных форм поведения, долго вырабатываемых и с трудом перестривающихся). А оно ведёт к самым разнообразным ошибкам, при применении к человеку просто опасно.
Источники
Cooper L., 1982. Internal representation// Animal mind-human mind. Report of the Dahlem Workshop on Animal Mind-Human Mind. Ed. D.Griffin. Berlin, Springer Verlag. 450 pp.
Hollard V., Delius J., 1983. Rotational invariance in visual pattern recognition by pigeons and humans// Science. Vol.218. P.804-806.
Lorenz K., 1937а. Űber die Bildung des Instinktbegriffes// Naturwiss. Bd.25. S.289-300, 307-318, 324-331.
Lorenz, K., 1937b. Űber den Begriff der Instinkthandlung // Folia Biotheoretica. Ser. B. Bd. 2. Instinctus. S.17-50.
via wolf_kitses
Организация инстинктивного поведения
Важный вопрос: под влиянием чего дятел, раздолбив шишку, летит за другой; срывает отточенным движением её с кормового дерева; несёт в клюве (реже в лапах) на кузницу, там прижимает грудью к стволу, после чего вынимает старую шишку из «станка» и выбрасывает, а потом ставит на её место новую и начинает долбить? Под влиянием голода? Отнюдь нет: исследования бюджета времени и энергии показывают, что питание семенами хвойных обеспечивает дятлу в среднем девятикратный перевес «прихода энергии над «расходом» (Бардин, 1982).
Остаётся даже на «активное бездельничанье», как и у других видов птиц и млекопитающих при устойчивой жизни, когда нет затяжной непогоды и других бедствий (см. его уморительное описание у самца серой вороны).
Так или иначе, видоспецифическое поведение здесь организуется прошлыми действиями, их успехом или неудачей, но отнюдь не голодом или иным побуждением (если дятла спугнуть при долблении, он летит на кормовое дерево, «автоматом» срывает шишку, но летит не на прежнюю кузницу, а в другую часть участка, где также есть кузница со старой, невытащенной шишкой, стимулирующей её заменить и «работать» над новой). До «толчка изнутри», от специфической (пищевой) мотивации, дело попросту не доходит, поведение опережает его. Сходным образом богач вносит деньги на счёт никак не для того, чтобы сберечь их от повседневных трат, а для последовательного увеличения свободы в будущих операциях, каковы бы они ни были.
Другой пример: хищническое поведение, особенно в отношении крупных жертв, способных эффективно сопротивляться, «спускается» отнюдь не голодом охотника, а беспомощностью (или, шире, уязвимостью большей чем обычно) добычи. Соответственно, оно не имеет других тормозов, кроме их исчерпания, когда все доступные жертвы мертвы или сумели отбиться. Если беспомощных жертв слишком много, хищник не останавливается, пока не зарежет всех, независимо от собственной сытости или усталости.
«Волки убивают всех находящихся в пределах досягаемости оленей или косуль, вязнущих в глубоком снегу, т.е. больше, чем могут съесть. Нет торможения реакции хищника при нападении на стадо домашних овец, в телятниках, либо на средних и малых копытных в вольерах и др.»
Вообще, для хищных млекопитающих показано, что одним из сигналов доступности жертвы, запускающих упорное преследование, являются изменения микрофлоры на поверхности мочки носа, свидетельствующие о долговременном стрессе, приведшем уже к какого- либо рода патологии. Практическим все животные, добытые хищником в природе, или же пойманные борзыми в специальных напусках на сайгаков и зайцев для изучения соответствующего феномена, несли разные серьёзные патологии. Напротив, охотники, стремившиеся отстрелять именно больных сайгаков или зайцев, примерно в трети случаев убивают вполне здоровых (Северцов, Шубкина, 2015).
Что будет, если ключевых раздражителей долго нет? Будут, как и считали классические этологи, «реакции вхолостую», «нулевой уровень изменчивости» инстинктивного поведения. Только организация их стохастическая, см. «Про смену парадигм в нейрофизиологии«. В.А.Непомнящим показано («Как животные решают плохо формализуемые задачи поиска«), что это в условиях неопределённости даёт наибольший шанс удовлетворить побуждение и/или в приемлемый срок вновь «натолкнуться» на действие ключевых раздражителей, организующих поведение закономерным и специфическим образом. На эндогенную детерминацию поведения места (и времени) снова не остаётся, она лишь источник устойчивости последнего при развёртывании.
Свежие комментарии