 Print
Print  PDF
PDF
Доместикация диких животных – краеугольный камень цивилизации. Идея Д.К. Беляева об основе доместикации как бессознательном отборе человеком каменного века неагрессивных животных была подтверждена многолетними уникальными работами Института цитологии и генетики по доместикации серебристо-черных лисиц, крыс-пасюков и американских норок. Обзор посвящен роли медиаторов мозга в механизмах доместикации. Полученные в наших исследованиях данные свидетельствуют об участии серотониновой системы мозга в перестройке поведения при селекции животных на низкую агрессивность. Установлено, что доместикация, превращающая агрессивное дикое животное в неагрессивное и дружелюбное, сопровождается у животных разных видов (серебристо-черных лисиц, крыс-пасюков и норок) изменением основных молекулярных элементов, определяющих функциональную активность серотониновой системы. Агрессивные и неагрессивные животные различаются по уровню серотонина в мозге, по активности основных ферментов метаболизма серотонина и по экспрессии серотониновых рецепторов. Активность катализирующей синтез серотонина в мозге триптофангидроксилазы-2 у неагрессивных животных повышена, активность фермента деградации моноаминоксидазы А – понижена.
Доместицированные крысы отличаются от агрессивных и по экспрессии серотониновых 5-НТ1А-рецепторов: уровень мРНК 5-НТ1А-рецептора, плотность рецепторов в структурах мозга и функциональная активность у них выше, чем у агрессивных. Все это свидетельствует о существенной роли нейромедиатора мозга серотонина в эволюционном процессе доместикации животных. Особое внимание в обзоре обращено на роль медиаторов мозга в коррелятивной изменчивости при селекции по поведению. Представлен комплекс доказательств нашей гипотезы, согласно которой одним из механизмов возникновения важнейших коррелятивных признаков при доместикации – изменений в основной системе стресса (гипоталамо-гипофизарно-надпочечниковой) и регулятора репродукции (гипоталамо-гипофизарно-половой) – является вовлечение в селекционный процесс серотониновой системы мозга, которая, наряду с функцией регулятора агрессивного поведения, контролирует эндокринные железы.
Доместикация диких животных – одно из величайших исторических событий, предопределивших развитие современной цивилизации. Ключевым признаком доместицированных животных является отсутствие агрессии по отношению к человеку. Идея академика Д.К. Беляева об основе доместикации как происходившем еще в глубокой древности бессознательном отборе человеком неагрессивных животных (Belyaev, 1979) была блестяще подтверждена многолетними уникальными работами Института цитологии и генетики по доместикации серебристо-черных лисиц (Trut, 1999), крыс-пасюков (Nikulina et al., 1986; Plyusnina, Oskina, 1997) и американских норок (Трапезов, 2007, 2008).
Несомненной заслугой Д.К. Беляева является последовательное развитие представлений о роли поведения и стресса в доместикации диких животных. Им были инициированы исследования нейрофизиологических и гормональных механизмов, которые и определяют особенности поведения и физиологических свойств животных, селекционируемых на низкую агрессивность по отношению к человеку.
Принципиальные программные положения о роли «регуляторов» поведения в доместикации животных сформулированы Д.К. Беляевым в начале 1960-х годов. Первая статья, в которой намечены основные идеи его теории доместикации и упомянуты регуляторы, была посвящена проблемам коррелятивной изменчивости и опубликована в «Известиях Сибирского отделения АН СССР» в 1962 г. (Беляев, 1962). В это же время он, ссылаясь на работы И.П. Павлова, писал:
«…стало ясно, что свойства основных нервных процессов играют весьма существенную роль в эволюционной судьбе особей и являются поэтому тем материалом, на основе которого действует естественный отбор» (Беляев, Трут, 1964).
Разрабатывая проблему коррелятивной изменчивости, Беляев высказал мысль о том, что сущность доместикации животных кроется в наследственно закрепляемом изменении регуляторных систем – гормональной и центральной нервной. Ориентация Д.К. Беляева на изучение нейрофизиологических и нейроэндокринных механизмов доместикации была плодотворной и в те годы, безусловно, новой.
В своих первых статьях Д.К. Беляев писал о нейрогуморальной регуляции и мозге вообще, без упоминания о медиаторах мозга. Это неудивительно, так как само существование в мозге медиаторных систем было установлено лишь к концу 1950-х годов, и их роль в регуляции поведения и физиологических функций во многом еще была неясна. Но постановка проблемы и ориентация исследователей на выявление тех процессов, которые реализуют действие отбора по поведению, на интеграцию теории искусственного отбора с современными представлениями о функциях мозга свидетельствует о научной интуиции и широте взглядов Д.К. Беляева.
Серотонин и доместикация
Нашими исследованиями (Попова и др., 1975; Popova et al., 1976, 1997; Popova, 1999) впервые показано, что доместикация животных сопровождается перестройкой нейромедиаторных систем мозга. Значительные изменения метаболизма и рецепции дофамина и важнейшего регулятора агрессивности – серотонина, были обнаружены у животных, селекционированных на отсутствие агрессивности по отношению к человеку. Особое внимание привлекают изменения в серотониновой (серотонергической) системе мозга. Это обусловлено рядом причин.
Во-первых, серотонин (5-гидрокситриптамин, 5-НТ) – эволюционно очень древний медиатор. Многочисленные данные свидетельствуют о его участии в регуляции агрессивного поведения не только млекопитающих (Попова и др., 1978; Valzelli et al., 1981; Molina et al., 1987; Miczek et al., 1989; Popova, 2006), но и животных других классов – птиц (Sperry et al., 2003), рыб (Adams et al., 1996), ракообразных (Huber, 2005). Во-вторых, клинические данные показывают ассоциацию импульсивной агрессивности человека и суицида, который рассматривают как проявление агрессии, направленной на самого себя (Pedder, 1992), с недостаточной функциональной активностью серотониновой системы мозга (Linnoila et al., 1983; Linnoila, Virkkunen, 1992; Arango et al., 2003). В-третьих, ранняя экспрессия в пренатальном периоде и выраженное влияние на морфогенез мозга определяют существенную роль серотонина в развитии мозга (Chubakov et al., 1986; Whitaker-Azmitia et al., 1996; Gaspar et al., 2003). И наконец, серотонин – наиболее экспансивный медиатор мозга, участвующий в регуляции не только поведения, но и ряда физиологических систем, включая важнейшую – нейроэндокринную систему (Naumenko, 1973; Науменко, Попова, 1975).
Функциональная активность серотониновой системы зависит от интенсивности синтеза и разрушения серотонина, обратного захвата (reuptake) и депонирования выделившегося в синаптическую щель серотонина, плотности серотониновых 5-НТ-рецепторов. Это определяет круг основных мишеней в генетической регуляции серотонином поведения – гены и ферменты, контролирующие синтез и разрушение серотонина в мозге, 5-НТ-транспортер и 5-НТ-рецепторы.
Синтезируется серотонин мозга из поступающей с пищей незаменимой аминокислоты триптофан. Катализируют его синтез два фермента – триптофангидроксилаза и декарбоксилаза ароматических аминокислот. Декарбоксилаза не является селективным для серотонина ферментом. Скорость его синтеза в мозге определяет специфичная для серотонина триптофангидроксилаза-2 (на периферии – триптофангидроксилаза-1). Синтезированный серотонин депонируется в нейронах. В ответ на нервный импульс он выделяется в синаптическую щель и действует на серотониновые 5-НТ-рецепторы, после чего инактивируется обратным захватом в те же нейроны. Этот замечательный процесс, происходящий с участием 5-НТ-транспортера, позволяет многократно использовать одну и ту же молекулу серотонина. Основной фермент деградации серотонина – моноаминоксидаза А (МАО А), основной конечный его метаболит – 5-гидроксииндолуксусная кислота.
Около 40 лет назад нами было впервые установлено, что доместикация связана с изменением в мозге метаболизма серотонина. Обнаружено (Попова и др., 1975), что уровень серотонина и его основного метаболита, 5-гидроксииндолуксусной кислоты, в ряде структур мозга доместицированных серебристо-черных лисиц существенно выше, чем у агрессивных животных. Далее было выяснено, что повышенный уровень серотонина в мозге лисиц, проявлявших дружелюбную реакцию на человека, связан с изменением экспрессии основных ферментов его метаболизма – повышением активности ключевого фермента синтеза серотонина триптофангидроксилазы-2 и понижением активности основного фермента его катаболизма – МАО А (Popova et al., 1991b, 1997). Позднее сходные изменения были обнаружены и у селекционируемых на низкую агрессивность крыс-пасюков. Уровень серотонина в среднем мозге и гипоталамусе, уровень его метаболита в гипоталамусе и активность триптофангидроксилазы неагрессивных крыс были повышены (Никулина и др., 1985а; Popova et al., 1991a).
Надо отметить, что различия в уровне серотонина в структурах мозга доместицированных и агрессивных крыс выявляются не всегда, более чувствительный показатель – уровень основного метаболита серотонина 5-гидроксииндолуксусной кислоты (Науменко и др., 1987). Отчетливые различия проявляются в реакции серотониновой системы на эмоциональный стресс: вызываемое стрессом снижение уровня серотонина в гипоталамусе и в среднем мозге агрессивных крыс было гораздо значительнее, чем у доместицируемых (Дыгало и др., 1985).
Участие серотонина мозга в регуляции агрессивного поведения и в механизмах доместикации было показано и на третьем виде животных – американской норке (Никулина и др., 1985б, 1993; Войтенко и др., 1992; Осадчук, Войтенко, 1992). Активность ферментов деградации серотонина – МАО А и МАО B – в стволе мозга неагрессивных норок оказалась пониженной. Введение предшественника серотонина, повышающего содержание этого медиатора в мозге, снижало агрессивность норок. Полифункциональность вездесущего серотонина связана с необычно высоким полиморфизмом его рецепторов.
Суперсемейство клонированных и секвенированных 5-НТ-рецепторов состоит из 15 подтипов, образующих 7 семейств. Эта классификация основана на трех основных характеристиках рецепторов: структуре (первичная последовательность аминокислот), характере передачи сигнала и реакции на фармакологические препараты. В последние годы наше внимание направлено главным образом на изучение роли 5-НТ1А-рецептора.
5-НТ1А-рецептор занимает особое место в обширном семействе 5-НТ-рецепторов. Он является основным ауторегулятором активности серотониновой системы мозга (Pineyro, Blier, 1999), участвует в регуляции уровня тревожности и депрессии, действуя как анксиолитик и антидепрессант (Nutt, Glue, 1991; Lucki et al., 1994), и понижает агонистическую и социальную активность крыс и мышей (Bell, Hobson, 1994; Olivier et al., 1995).
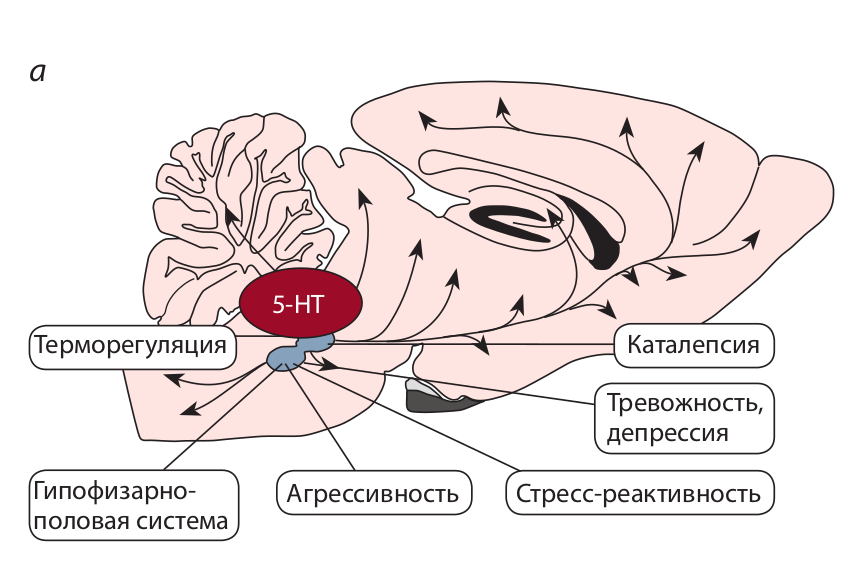
Серотонин (5-НТ) мозга и коррелятивные признаки доместикации. а – полифункциональность серотонина мозга; б – влияние селекции по
агрессивному поведению на серотониновую систему, агрессивность и серотонин-зависимые нейроэндокринные системы – гипоталамо-гипофизарно-надпочечниковую (стрессорная реактивность) и гипоталамо-гипофизарно-половую.
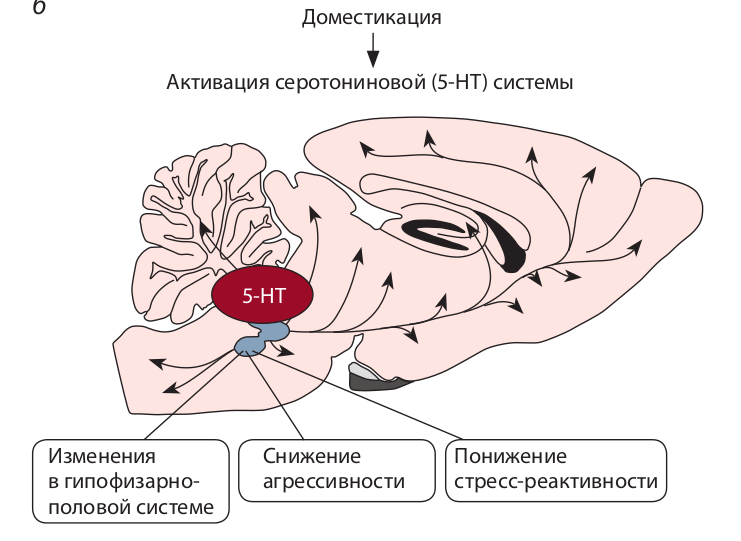
Было показано (Popova et al., 2005), что доместицированные крысы значительно отличаются от агрессивных по экспрессии 5-НТ1А-рецепторов: уровень мРНК 5-НТ1А-рецептора, плотность рецепторов в структурах мозга и функциональная активность у них выше, чем у
агрессивных крыс. Полученные данные свидетельствуют о том, что доместикация, превращающая агрессивное дикое животное в совершенно неагрессивное и дружелюбное по отношению к человеку, сопровождается радикальным изменением всех основных элементов, определяющих функциональную активность серотониновой системы. Принципиально важно, что изменения серотониновой системы мозга обнаружены при доместикации разных, достаточно далеких видов млекопитающих – серебристо-черных лисиц, крыс-пасюков и норок. Это говорит о существенной роли нейромедиатора мозга серотонина в эволюционном процессе доместикации животных и хорошо соответствует сложившемуся представлению о серотонине как о факторе, понижающем агрессивность большинства видов животных.
Серотонин и коррелятивная изменчивость
Селекция животных нередко сопровождается появлением коррелятивных признаков, т. е. признаков, по которым селекция не велась. Особенно многочисленны и часто неожиданны коррелятивные признаки при селекции по поведению (Беляев, 1962; Belyaev, 1969). Многообразные морфологические и физиологические изменения, характерные для доместицированных животных, детально описаны Л.Н. Трут (Trut, 1999). Несмотря на то что проблема коррелятивной изменчивости со времен Дарвина вызывает пристальный интерес, механизмы появления многообразных коррелятивных признаков все еще во многом неясны. Поразительно то, что при селекции по доместикационному типу поведения многие из них аналогичны у разных видов животных (Trut, 1999). Эта загадочное свойство селекции по поведению, несомненно, свидетельствует о сходных механизмах коррелятивной изменчивости.
Доказательства существенных сдвигов, происходящих в процессе доместикации в одной из основных систем стресса – гипоталамо-гипофизарно-надпочечниковой – и в функциональной активности гонад, были получены более 40 лет тому назад исследованиями лаборатории проф. Е.В. Науменко (Науменко и др., 1971, 1974; Трут и др., 1972). Установлено, что у доместицируемых серебристо-черных лисиц уровень в крови гормонов коры надпочечников во все сезоны года, кроме периода размножения, ниже, чем у неселекционируемых лисиц. Различна и реактивность центральных механизмов регуляции гипоталамо-гипофизарно-надпочечниковой системы, что проявляется в разной реакции на психический стресс (Трут и др., 1972).
Появление сходных различий было обнаружено и в ходе селекции на доместикационный тип поведения другого вида – крыс (Naumenko et al., 1989). Крысы-пасюки, селекционируемые на отсутствие агрессивной реакции на человека, отличались от высокоагрессивных крыс не только кротким и даже дружественным поведением по отношению к человеку, но и пониженным уровнем кортикостерона в крови и пониженной реакцией гипоталамо-гипофизарно-надпочечниковой системы на эмоциональный стресс и на центральное введение (в латеральный желудочек мозга) серотонина (Дыгало и др., 1985).
Селекция на доместикационный тип поведения значительно изменяет эндокринную регуляцию половой системы и репродуктивность животных. У «ручных» серебристо-черных лисиц:
1) изменена фотопериодическая регуляция эндокринной функции яичников: размножаются они в тот же сезон, но раньше, чем лисицы дикого типа поведения;
2) потенциальная плодовитость выше, чем у агрессивных, но выше и эмбриональная смертность;
3) у самцов сезонное угасание эндокринной функции семенников начинается раньше (Осадчук и др., 1978; Осадчук, Трут, 1989; Осадчук, 2006).
Как уже было отмечено, серотонин считается одним из наиболее экспансивных нейромедиаторов мозга. Он регулирует не только поведение и эмоциональность, но и такие физиологические функции, как терморегуляция, сон, зимняя спячка зимоспящих, суточные ритмы, пищевое поведение. Важнейшей его ролью является участие в регуляции гипоталамо-гипофизарно-надпочечниковой и гипоталамо-гипофизарно-половой систем (Науменко, Попова, 1975).
Значение серотонина мозга в регуляции гипоталамо-гипофизарно-надпочечниковой системы показано давно (Naumenko, 1973) и не вызывает сомнения (Jorgensen, 2007). Серотонин считается медиатором, тесно связанным с реакцией на стресс (stress-related neurotransmitter) (Yamaguchi et al., 2016). Накоплены многочисленные данные, свидетельствующие и об участии серотонина мозга в регуляции половых гормонов и в половой мотивации самцов (Науменко и др., 1983; Popova, Amstislavskaya, 2002).
Полифункциональность серотонина, его признанная роль основного центрального регулятора агрессивности и выявленные нашими исследованиями значительные изменения в серотониновой системе мозга у разных видов доместицируемых животных дали нам основание предположить, что причиной появления ряда коррелятивных признаков является изменение этой медиаторной системы. К таким коррелятивным признакам относятся и важнейшие – изменение гормональной регуляции гипофизарно-половой системы, изменение функциональной активности гипофизарно-надпочечникой системы и связанное с этим понижение реакции на стресс (Попова и др., 1980) (рисунок).
Эта гипотеза объясняет две загадочные особенности селекции по поведению – частое возникновение коррелятивных признаков и появление сходных коррелятивных признаков при доместикации разных видов животных (Беляев, 1962; Belyaev, 1969; Trut, 1999). Она же наполняет конкретным содержанием использовавшееся ранее объяснение – плейотропное действие гена. Медиаторы мозга – основные регуляторы поведения, и каждый вид поведения контролируется «ансамблем» нейромедиаторов, в котором есть один или ограниченное число «ведущих солистов». Селекция по поведению по своей сути является селекцией на определенную функциональную активность основной из контролирующих данный вид поведения медиаторных систем. Все медиаторы мозга в той или иной степени полифункциональны, и изменение метаболизма или рецепции медиатора не только ведет к изменению поведения, но и влияет на регуляцию других контролируемых этим медиатором физиологических систем. Это и определяет частое появление коррелятивных признаков при селекции по поведению.
Сходные коррелятивные признаки при доместикации разных видов животных обусловлены сходством нейромедиаторных механизмов, регулирующих агрессивность. Серотонин регулирует не только агрессивное поведение всех изученных млекопитающих, но и, как уже упоминалось, поведение животных других классов.
Разумеется, изменением функциональной активности серотониновой системы невозможно объяснить весь спектр коррелятивных признаков, возникающих при доместикации животных. Серотонин – это ключевой медиатор (key player) в механизмах агрессивности, но, несомненно, не единственный. Это слишком сложный эксперимент природы, в котором участвуют и другие медиаторы, прежде всего дофамин и норадреналин (Дыгало и др., 1985; Дыгало, 1986; Никулина, 1990). Так, показано, что в структурах мозга крыс, селекционируемых на доместикационное поведение, изменена активность тирозингидроксилазы – фермента синтеза дофамина и норадреналина. Есть основания предполагать, что в изменении стрессорной реактивности доместицируемых крыс наряду с серотониновой системой существенную роль играет норадренергическая система мозга (Дыгало, 1986; Науменко и др., 1987; Дыгало и др., 1988). Недавно нами обнаружено (Ilchibaeva et al., 2015, 2016), что неагрессивные крысы отличаются от высокоагрессивных и по экспрессии нейротрофических факторов – BDNF (brain derived neurotrophic factor) и GDNF (glial cell derived neurotrophic factor). Нейротрофические факторы, не являясь медиаторами, играют важнейшую роль, регулируя рост и развитие нейронов и синаптических связей мозга, и наравне с серотониновой системой представляют важнейшие факторы нейропластичности.
В организме все соединено многообразными связями, и до полного понимания всей этой сложнейшей перестройки мозга еще далеко, хотя уже то, что стало известно, говорит о глубочайших различиях мозга агрессивного и неагрессивного фенотипа. Кроме проблемы доместикации, эти данные относятся к еще одной проблеме – механизмам генетической предрасположенности к агрессивному поведению, которая, к сожалению, становится все более актуальной.
В отношении серотониновой системы пока остаются открытыми многие вопросы. Среди них вопрос, мутация какого гена является первичной, а каких – происходит в результате последовавших адаптивных изменений мозга. Проблема осложняется тем, что изменение экспрессии любого из ключевых генов, контролирующих серотониновую систему, нарушает ее функционирование и может повлиять на агрессивность. Так, более агрессивны мыши с нокаутом МАО А (Cases et al., 1995; Popova et al., 2001) и мужчины – носители однолокусной мутации МАО А (Brunner, 1995). Нокаут гена, контролирующего транспортер серотонина, понижает агрессивность мышей (Holmes et al., 2002). Влияет на агрессивность и изменение экспрессии гена триптофангидроксилазы-2, контролирующего синтез серотонина. Показано, что C1473G полиморфизм гена триптофангидроксилазы-2 влияет не только на скорость синтеза серотонина, но и на агрессивность мышей (Zhang et al., 2004; Kulikov et al., 2005). Трудно представить, что одновременно возникли мутации во всех ключевых генах серотониновой системы. Скорее всего, первоначально это была мутация какого-то одного из них, а затем, в процессе селекции на отсутствие агрессивности по отношению к человеку, произошла перестройка всей серотониновой системы мозга доместицированного животного. Это стало также причиной появления некоторых коррелятивных признаков.
Фотоальбом к 40-летию ИЦиГ СО РАН
Время все расставляет по своим местам. Представления Д.К. Беляева привлекли нейрофизиологов и нейроэндокринологов и привели к появлению в Институте цитологии и генетики СО АН нового направления – физиологической генетики, ставшей предшественником современной нейрогеномики. И теперь уже можно уверенно применить определение «впервые» к экспериментально подтвержденным в оригинальных исследованиях ИЦиГ СО АН (РАН) идеям Д.К. Беляева о роли поведения, его регуляторов и стресса в эволюционном процессе доместикации животных. Впервые было обнаружено влияние селекции на неагрессивное поведение на гипоталамогипофизарно-надпочечниковую и половую нейроэндокринные системы. Впервые было выявлено участие в механизмах доместикации медиаторов мозга, прежде всего серотонина, и сформулировано представление о его роли в возникновении коррелятивных признаков. Вклад Дмитрия Константиновича Беляева в обоснование эволюционной связи доместикации и поведения животных и в развитие направления, которое вошло в ХХI век под названием нейрогеномики и считается одним из наиболее актуальных и перспективных направлений современной генетики, несомненен.
References
Adams C.F., Liley N.R., Gorzalka B.D. PCPA increases aggression in in male firemouth cichlids. Pharmacology. 1996;53:328-330.
Arango V., Huang Y., Underwood M.D., Mann J.J. Genetics of the serotonin system in suicidal behavior. J. Psychiat. Res. 2003;37:375-386.
Bell R., Hobson H. 5-HT 1A receptor influences on rodent social and agonistic behavior. Neurosci. Biobehav. Rev. 1994;18:325-338.
Belyaev D.K. Problems of correlative alterations and their significance for the theory of evolution and for animal breeding. Izvestiya SO AN SSSR = Proceedings of the Siberian Branch of the USSR Academy of Sciences. 1962;10:111-124. (in Russian)
Belyaev D.K. Domestication of animals. Sci. J. (UK). 1969;5:47-52.
Belyaev D.K. Destabilizing selection as a factor of domestication. J. Heredity. 1979;70:301-308.
Belyaev D.K., Trut L.N. Behavior and breeding of animals. II. Correlative changes under selection for tame behavior. Byulleten Moskovskogo obshchestva ispytateley prirody = Bulletin of Moscow
Society of Naturalists. 1964;4:5-9. (in Russian)
Brunner H.G. Monoamine oxidase and behavior. Ann. Med. 1995;27: 431-432.
Cases O., Seif I., Grimsby J., Gaspar P., Chen K., Pournin S., Müller U., Aguet M., Babinet C., Shih J.C., De Maeyer E. Aggressive behavior and altered amounts of brain serotonin and norepinephrine in mice lacking MAOA. Science. 1995;268:1763-1766.
Chubakov A.R., Gromova E.A., Konovalov G., Sarkisova F., Chumazov E. The effects of serotonin on morphofunctional development of rat cerebral neocortex in tissue culture. Brain Res. 1986;389:
285-297.
Dygalo N.N. Role of the noradreneregic system of the brain in the transformation of stressor reactivity in Norway rats selected for domestic behavior. Zhurnal obshchey biologii = Journal of General Biology. 1986;4:455-458. (in Russian)
Dygalo N.N., Bykova T.S., Naumenko E.V. Tyrosine hydroxylase activity in the brain of silver foxes selected for the type of behavior. Zhurnal evolyutsionnoy biokhimii i fiziologii = Journal of Evolutionary Biochemistry and Physiology. 1988;24(4):503-507. (in Russian)
Dygalo N.N., Shishkina G.T., Borodin P.M., Naumenko E.V. The role of neurochemical systems of the brain in changes of reactivity of the pituitary-adrenocortical complex in Norway rats selected for behavior. Zhurnal evolyutsionnoy biokhimii i fiziologii = Journal of Evolutionary Biochemistry and Physiology. 1985;21(4):342-347. (in Russian)
Gaspar P., Cases O., Maroteaux L. The developmental role of serotonin: news from mouse molecular genetics. Nat. Rev. Neurosci. 2003; 4(12):1002-1012.
Holmes A., Murphy D., Crawley J. Reduced aggression in mice lacking the serotonin transporter. Psychopharmacology. 2002;161:160-167.
Huber R. Amines and motivated behaviors: a simpler systems approach to complex behavioral phenomena. J. Comp. Physiol. A. Neuroethol. Sens. Neural Behav. Physiol. 2005;191:231-239.
Ilchibaeva T.V., Kondaurova E.M., Tsybko A.S., Kozhemyakina R.V., Popova N.K., Naumenko V.S. Brain-derived neurotrophic factor (BDNF) and its precursor (proBDNF) in genetically defined fearinduced aggression. Behav. Brain Res. 2015;290:45-50.
Ilchibaeva T.V., Tsybko A.S., Kozhemyakina R.V., Popova N.K., Naumenko V.S. Glial cell line-derived neurotrophic factor in genetically defined fear-induced aggression. Eur. J. Neurosci. 2016;44(7):2467-2473.
Jorgensen H.S. Studies of neuroendocrine role of serotonin. Dan. Med. Bull. 2007;54(4):266-288.
Kulikov A.V., Osipova D.V., Naumenko V.S., Popova N.K. Association between Tph2 gene polymorphism, brain tryptophan hydroxylase activity and aggressiveness in mouse strains. Genes Brain Behav. 2005;4(8):482-485.
Linnoila V.M., Virkkunen M. Aggression, suicidality, and serotonin. J. Clin. Psychiatry. 1992;53(10):46-51.
Linnoila V.M., Virkkunen M., Schwannian M., Nuutila A., Rimon R., Goodwin F.K. Low cerebrospinal fluid 5-hydroxyindoleacetic acid concentration differentiates impulsive from non-impulsive violent behavior. Life Sci. 1983;33:2609-2614.
Lucki I., Singh A., Kreiss D.S. Antidepressant-like behavioral effects of serotonin receptor agonists. Neurosci. Biobehav. Rews. 1994;18(1): 85-95.
Miczek K.A., Mos J., Olivier B. Brain 5-HT and inhibition of aggressive behavior in animals: 5-HIAA and receptor subtypes. Psychopharmacol. Bull. 1989;25:399-403.
Molina V., Ciesielski L., Gobaille S., Isel F., Mandel P. Inhibition of mouse-killing behavior by serotonin-mimetic drugs: effects of partial alterations of serotonin neurotransmission. Pharmacol. Biochem. Behav. 1987;27:123-131.
Naumenko E.V. Central regulation of the pituitary-adrenal complex. N. Y.: Plenum Publ. Corp., 1973.
Naumenko E.V., Osadchuk A.V., Serova L.I., Shishkina G.T. Genetikofiziologicheskie mekhanizmy regulyatsii funktsiy semennikov [Genetic and physiological mechanisms in testis regulation]. Novosibirsk: Nauka Publ., 1983;203. (in Russian)
Naumenko E.V., Popova N.K. Serotonin i melatonin v regulyatsii endokrinnoy sistemy [Serotonin and melatonin in the regulation of endocrines]. Novosibirsk: Nauka Publ., 1975;218. (in Russian)
Naumenko E.V., Popova N.K., Ivanova L.N. Neuroendocrine and neurochemical mechanisms of domestication. Genetika = Genetics (Moscow). 1987;23(6):1011-1026. (in Russian)
Naumenko E.V., Popova N.K., Nikulina E.M., Dygalo N.N., Shishkina G.T., Borodin P.M., Markel A.L. Behavior, adrenocortical activity, and brain monoamines in Norway rats selected for reduced aggressiveness towards man. Pharmacol. Biochem. Behav. 1989;33:85-91.
Naumenko E.V., Trut L.N., Korshunov E.A., Belyaev D.K. Functioning of adrenal cortex and its seasonal changes in silver foxes. Doklady Akademii nauk SSSR = Reports of the Academy of Sciences of USSR. 1971;200(5):1249-1251. (in Russian)
Naumenko E.V., Trut L.N., Pavlova S.I., Belyaev D.K. Genetics and phenogenetics of hormonal traits in animals: 1. Functional correlation between adrenal and gonad glands in silver foxes and changes of the correlation under selection for behavior. Genetika = Genetics (Moscow). 1974;10(10):52-57. (in Russian)
Nikulina E.M. Brain catecholamines in domestication of silver foxes Vulpes fulvus. Zhurnal evolyutsionnoy biokhimii i fiziologii = Journal of Evolutionary Biochemistry and Physiology. 1990; 26(2):156-160. (in Russian)
Nikulina E.M., Borodin P.M., Popova N.K. Changes in certain forms of aggressive behavior and brain monoamine levels during the selection of wild rats for tame behavior. Zhurnal vysshey nervnoy deyatelnosti im. I.P. Pavlova = I.P. Pavlov Journal of Higher Nervous Activity. 1985а;35(4):703-709. (in Russian)
Nikulina E.M., Borodin P.M., Popova N.K. Changes in certain form of aggressive behavior and monoamine content in the brain during selection of wild rats. Neurosci. Behav. Physiol. 1986;16(6):
466-471.
Nikulina E.M., Trapezov O.V., Kharlamova A.A., Popova N.K. Affective defensive behavior in minks: influence on serotonergic and noradrenergic brain systems. Sibirskiy biologicheskiy zhurnal = Siberian Journal of Biology. 1993;3:9-12. (in Russian)
Nikulina E.M., Trapezov O.V., Popova N.K. Monoamine contents in the brain of minks differing in the response to man. Zhurnal vysshey nervnoy deyatelnosti im. I.P. Pavlova = I.P. Pavlov Journal of Higher Nervous Activity. 1985b;35(6):1142-1146. (in Russian)
Nutt D.J., Glue P. Clinical pharmacology of anxiolytics and antidepressants: a psychopharmacological perspective. Psychopharmacology of Anxiolytics and Antidepressants. Ed. S.E. File. N. Y: Perg. Press, 1991:1-28.
Olivier B., Mos J., van Oorschot R., Hen R. Serotonin receptors and animal models of aggressive behavior. Pharmacopsychiat. 1995; 28:80-90.
Osadchuk L.V. Reproductive potential in male silver foxes under long-lasting selection for domesticated behavior. Zhurnal evolyutsionnoy biokhimii i fiziologii = Journal of Evolutionary Biochemistry and Physiology. 2006;42(2):146-152. (in Russian)
Osadchuk L.V., Krass P.V., Trut L.N., Ivanova L.N. Gonadal endocrine function in male silver foxes with different hereditary determined forms of defensive behavior. Izvestiya SO AN SSSR. Seriya biologicheskikh nauk = Proceedings of the Siberian Branch of the USSR Academy of Sciences. Biological series. 1978;2:79-85. (in Russian)
Osadchuk L.V., Trut L.N. Endocrine function of gonads in female silver foxes selected for domesticated behavior. Zhurnal obshchey biologii = Journal of General Biology. 1989;50(2):189-198. (in Russian)
Osadchuk L.V., Voitenko N.N. Sex steroids and brain serotonin in silver foxes during the estrus cycle. Rossiyskiy fiziologicheskiy zhurnal im. I.M. Sechenova = I.M. Sechenov Physiological Journal. 1992;78(4):118-122. (in Russian)
Pedder J. Psychoanalytic views of aggression: some theoretical problems. Br. J. Med. Psychol. 1992;65:95-106.
Pineyro G., Blier P. Autoregulation of serotonin neurons: role in antidepressant drug action. Pharmacol. Rews. 1999;51(3):533-591.
Plyusnina I., Oskina I. Behavioral and adrenocortical responses to open-field test in rats selected for reduced aggressiveness toward humans. Physiol. Behav. 1997;61:381-385.
Popova N.K. Brain serotonin in genetically defined defensive behavior.
Complex Brain Functions: Conceptual Advances in Russian Neuroscience. Eds. R. Millar, A.M. Ivanitsky, P.M. Balaban. N. Y.: Harwood Press, 1999;317-329.
Popova N.K. From genes to aggressive behavior: the role of serotonergic system. BioEssays. 2006; 28(5):495-503.
Popova N.K., Amstislavskaya T.G. Involvement of the 5-HT1A and 5-HT1B receptor sutypes in sexual arousal in male mice. Psychoneuroendocrinology. 2002;27:609-618.
Popova N.K., Gilinsky M.A., Amstislavskaya T.G., Morosova E.A., Seif I., De Maeyer E. Regional 5-HT metabolism in the brain of transgenic mice lacking monoamine oxidase A. J. Neurosci. Res.
2001;(66):423-427.
Popova N.K., Kulikov A.V., Avgustinovich D.F., Voitenko N.N., Trut L.N. Effect of domestication on the basic enzymes of serotonin metabolism and serotonin receptors in silver foxes. Rus. J. Genet.
1997;33(3):370-374.
Popova N.K., Kulikov A.V., Nikulina E.M., Kozlachkova E.Y., Maslova G.B. Serotonin metabolism and serotonergic receptors in Norway rats selected for low aggressiveness to man. Aggr. Behav. 1991A; 17:207-213.
Popova N.K., Naumenko E.V., Kolpakov V.G. Serotonin i povedenie [Serotonin and behavior]. Novosibirsk: Nauka Publ., 1978;304. (in Russian)
Popova N.K., Naumenko V.S., Plyusnina I.Z., Kulikov A.V. Reduction in 5-HT 1A receptor density, 5-HT 1A mRNA expression, and functional correlates for 5-HT 1A receptors in genetically defined aggressive rats. J. Neurosci. Res. 2005;80:286-292.
Popova N.K., Voitenko N.N., Kulikov A.V., Avgustinovich D.F. Evidence for the involvement of central serotonin in mechanism of domestication of silver foxes. Pharmacol. Biochem. Behav. 1991B;40: 751-756.
Popova N.K., Voitenko N.N., Pavlova S.I., Trut L.N., Naumenko E.V., Belyaev D.K. Genetics and phenogenetics of hormonal characteristics in animals: VII. Functional correlation between brain serotonin and the hypothalamus-pituitary-adrenal axis under emotional stress in domesticated and wild-type silver foxes. Genetika = Genetics (Moscow). 1980;16(10):1865-1871. (in Russian)
Popova N.K., Voitenko N.N., Trut L.N. Changes in the serotonin and 5-hydroxyindoleacetic acid brain levels in silver foxes selected for behavior. Doklady Akademii nauk SSSR = Reports of the Academy of Sciences of USSR. 1975;2(33):498-500. (in Russian)
Popova N.K., Voitenko N.N., Trut L.N. Changes in the content of serotonin and 5-hydroxyidoleacetic acid in the brain in the selection of silver foxes according to behavior. Neurosci. Behav. Physiol. 1976; 7:72-74.
Sperry T.S., Thomson C.K., Wingfield J.C. Effects of acute treatment with 8-OH-DPAT and fluoxetine on aggressive behavior in male song sparrows (Melospiza melodia morphna). J. Neuroendocrinol. 2003;15:150-160.
Trapezov O.V. On fur animal domestication (on the 140-year anniversary of Charles Darwin’s work “The variation of plants and animals under domestication”, published in Russia). Informatsionnyy vestnik VOGiS = The Herald of Vavilov Society for Geneticists and Breeders. 2007;11(1):45-61. (in Russian)
Trapezov O.V. The regulation role of genes of behavior in coat color formation in American mink (Mustela vison Schreber, 1777). Informatsionnyy vestnik VOGiS = The Herald of Vavilov Society for Geneticists and Breeders. 2008;12(1/2):63-83. (in Russian)
Trut L.N. Early canid domestication: the farm-fox experiment. Am. Scientist. 1999;87:160-169.
Trut L.N., Naumenko E.V., Belyaev D.K. Changes in the pituitaryadrenal function of silver foxes under selection for domestication. Genetika = Genetics (Moscow). 1972;8(5):35-43. (in Russian)
Valzelli L., Bernasconi S., Garattini S. p-Chlorophenylalanine-induced muricidal aggression in male and female laboratory rats. Neuropsychobiol. 1981;7:315-320.
Voitenko N.N., Maslova L.N., Trapezov O.V., Harlamova A.A. Brain monoamine oxidase in minks bred for response to humans. Zhurnal vysshey nervnoy deyatelnosti im. I.P. Pavlova = I.P. Pavlov Journal of Higher Nervous Activity. 1992;42(3):537-542. (in Russian)
Whitaker-Azmitia P.M., Druse M., Walker P., Lauder J.M. Serotonin as developmental signal. Behav. Brain Res.1996;73:19-39.
Yamaguchi N., Nakajima N., Ocada S., Yuri K. Effect of aging on stress-related responses of serotonergic neurons in the dorsalraphe nucleus of male rats. Neurobiol. Stress. 2016;3:43-51.
Zhang X., Beaulieu J.M., Sotnikova T.D., Gainetdinov R.R., Caron M.G. TPH-2 controls brain serotonin synthesis. Science. 2004; 305:217.
Вавиловский журнал генетики и селекции. 2017. Т.21. №4. С.414-420
Свежие комментарии