 Print
Print  PDF
PDF
По морфоэкологическому облику дятлы делятся на три большие группы — земляные дятлы (Erdspechte), долбящие дятлы (Hackspechte) и желны (Großspechte, крупные дятлы): это не филогенетические ветви, но уровни организации, выделенные по роли долбления и характеру лазания. См.подробней «20 фактов о дятлах» (п.18)
По морфоэкологическому облику дятлы делятся на три большие группы — земляные дятлы (Erdspechte), долбящие дятлы (Hackspechte) и желны (Großspechte, крупные дятлы): это не филогенетические ветви, но уровни организации, выделенные по роли долбления и характеру лазания. См.подробней «20 фактов о дятлах»
Доклад на семинаре ЦЭПЛ РАН 22.01.2021 г.
Резюме. Современная лесная парадигма, или gap-парадигма, описывает устойчивость лесных сообществ как динамический процесс (баланс между образованием «окон» или «прорывов» лесного полога и восстановлением его элементов в ходе популяционной жизни деревьев и кустарников). Их пространственная структура складывается из совокупности нарушений, находящихся на разных стадиях «зарастания» (так называемая мозаично-циклическая концепция организации экосистем). В развитие обоих реконструируется роль дятловых птиц в такого рода динамике, в зависимости от видов лесных биомов и степени их нарушенности. Показано, что
А) биоценотическая роль дятлов несводима к обычно упоминаемым «поставщикам дупел» и «санитарам леса»: то и другое — производные главного, участия этих птиц в воспроизводстве мозаичной структуры лесного полога, с характерной для данного биома архитектурой крон и составом деревьев-средообразователей 1-го яруса (вертикальное и горизонтальное измерения сложности организации лесного полога).
Благодаря этому дятлы участвуют в «посмертии дерева»: плотность подолбов разной площади и глубины, оставшихся от кормодобывания разных видов дятлов, поддерживает скорость разложения мёртвой древесины, оптимальную для данного лесного ценоза.
И наоборот: устойчивость обитания дятлов на данной территории зависит от обилия мёртвой древесины, особенно у специализированных видов — белоспинного Dendrocopos leucotos, трёхпалого Picoides tridactylus, среднего D.medius, или их американских аналогов — P.arcticus, P.tridactylus dorsalis, P.t.bacatus, D.albolarvatus.
Б) Для её осуществления необходимо всё биоразнообразие дятлов, не просто отдельные виды, чем большее число видов дятлов совместно обитает на одной территории, тем сложнее организация лесного полога, воспроизводимая их деятельностью, почему биоразнообразие дятловых растёт с увеличением натуральности лесного массива, а в ряду разных типов лесных биомов — от бореальных лесов к влажным тропическим.
В) В своей биоценотической роли дятлы действуют сопряжённо с дереворазрушающими грибами: они вектор их переноса при долблении. И наоборот: дятлы используют сделанные ими разрушения древесины как сигналы для «запуска» гнездостроения или выбора кормового метода, адекватного данному субстрату. Поражение ствола бактериальной и/или грибной инфекцией, в диапазоне от размягчения до образования полостей во многом — условие успешного осуществления того и другого и/или предпочтительно для дятлов.
Обсуждается индикаторная роль дятлов (биоразнообразия группы в целом и присутствия отдельных видов, особенно специализированных) в лесных биогеоценозах, показываются её нарушения в связи с урбанизацией, «освобождающей» те или иные виды дятлов (даже специализированные) от биоценотических органичений, заданных А-В).
Почему дятлов считают «ключевыми видами»?
Содержание
Настоящих дятлов п/сем. Picinae1 традиционно считают ключевыми видами лесных сообществ, англ. key species, или экосистемными инженерами (ecosystem engineers) [1]. Чаще всего это делается «за» две функции:
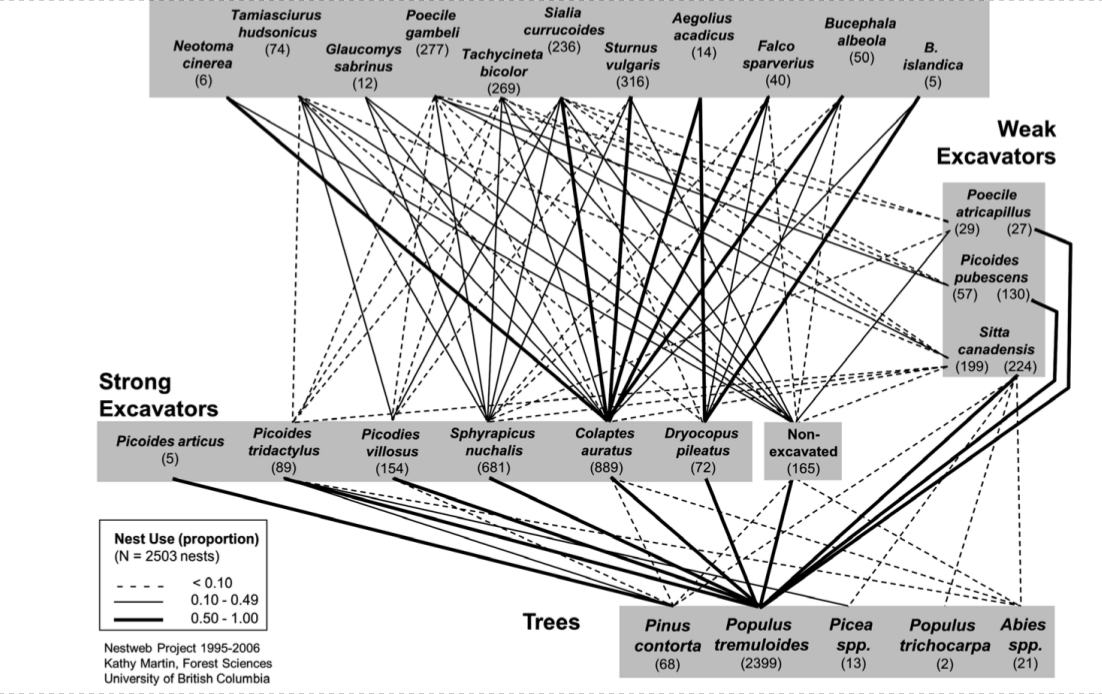
1) активное долбление дупел, гнёздовых и ночёвочных (владельцы внегнездовых территорий). Строители дупел чаще всего гнездятся там однократно (или ночуют только одну осень-зиму, даже когда этот участок занят круглогодично), хотя и повторное используют регулярно, и не только малодолбящие виды [2-4], но и самые специализированные к долблению (трёхпалый P.tridactylus) [5]. Возникает «фонд» пустующих дупел2: несколько следующих лет (4-7, максимально до 10 и 30) их используют «потребители», долбить неспособные или долбящие мало и плохо — разные виды птиц-вторичных дуплогнёздников, млекопитающих, общественных насекомых, в тропиках амфибий и рептилий [4, 6]. Они занимаются также другими дятлами, включая «доделку» и «укрупнение» малодолбящими формами готовых дупел более мелких видов, как зелёный Picus viridis — большого пёстрого дятла, золотой Colaptes auratus — волосатого D.villosus [7]. Виды, гнездящиеся в дятловых дуплах, часто считают зависимыми от присутствия видов, их продуцирующих: помимо дятлов, это выщипывающие гнёзда в гнилой древесине синицы-гаички Poecile spp., гренадёрки Lophophanes cristatus в умеренных широтах, трогоны Trogonidae и азиатские бородастики Megalaimidae в тропиках). Совокупность тех и других обозначают как «гнездовую сеть» (англ.nest web): см.примеры «сетей» Центра Нечерноземья [3], Беловежской Пущи [4], Южного Приморья [8], Британской Колумбии [9], Атлантического леса в Аргентине [10] (рис.1).
2) роль «санитаров леса» – пресечение вспышек массового размножения «вредителей»: разных видов насекомых-фитофагов, в том числе ксилофагов (короедов, лубоедов, заболонников, древоточцев, златок и усачей, муравьёв-древоточцев), голых гусениц и других листогрызущих, тлей и т.д. [2, 11-12]. Она распадается на две части.
А. Внутренние районы Британской Колумбии. Описывает создание дупел в разных видах деревьев выдалбливателями, «сильными» (большинство дятлов) и «слабыми» (черноголовая гаичка Parus atricapillus, пушистопёрый дятел Dendrocopos pubescens, каролинский поползень Sitta carolinensis), а также использование этого ресурса разными видами вторичных дуплогнёздников и других видов. Толщина и вид линий (слева внизу) пропорциональна частоте «производства» и «потребления» данными видами дупел в данных деревьях в изученном лесном массиве. Источник. Fig. 4.1 в: Mikusiński Grz., Roberge J.-M., Fuller R.J., 2018. Ecology and conservation of forest birds.
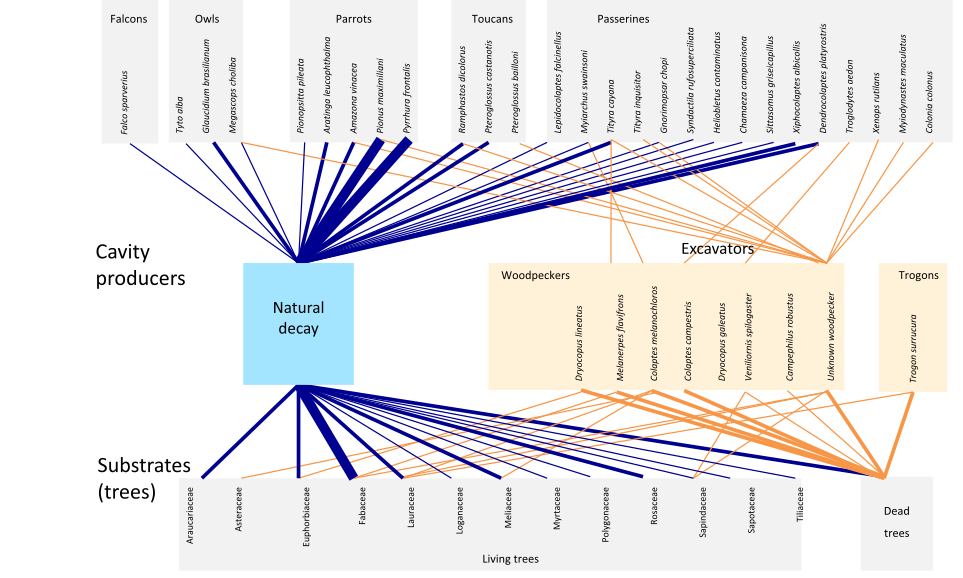
Б. «Атлантический лес» на юго-востоке Бразилии — одно из пяти важнейших «горячих точек» биоразнообразия (biodiversity hotspots) на планете (с включением более южных лесов Парагвая и аргентинской провинции Misiones, флористически и физиономических относящихся к тому же типу — именно их изучали цитируемые авторы). Влажные тропические леса здесь сильно сокращены человеком (осталось лишь 7%), оставшиеся фрагментированы (это «архипелаг», и сильно раздробленный, не «материк», как амазонская или оринокская сельва). Здесь крайне высок эндемизм, среди них много глобально-угрожаемых, из Красной книги МСОП, почему крайне важно эти леса сохранить.
Обозначения. Верхний ряд: «пользователи» дупел, сами их не выдалбливающие (попугаи, совы, туканы, соколы, воробьиные птицы). средний — источники дупел: естественное выгнивание и виды-»производители»: разные виды дятлов, 1 — трогонов, нижний — субстраты, в которых возникают дупла (живые деревья и сухостой). Линии — выбор и предпочтение, их толщина соответствует частоте случаев: тонкие 1-2, средние 3-4, толстые 5-6.
Источник. Fig.2 в: Cockle K.L., Martin K., Robledo G. Linking fungi, trees, and hole-using birds in a Neotropical tree-cavity network: Pathways of cavity production and implications for conservation// Forest Ecology and Management. 2012. V.264. Р.210-219.
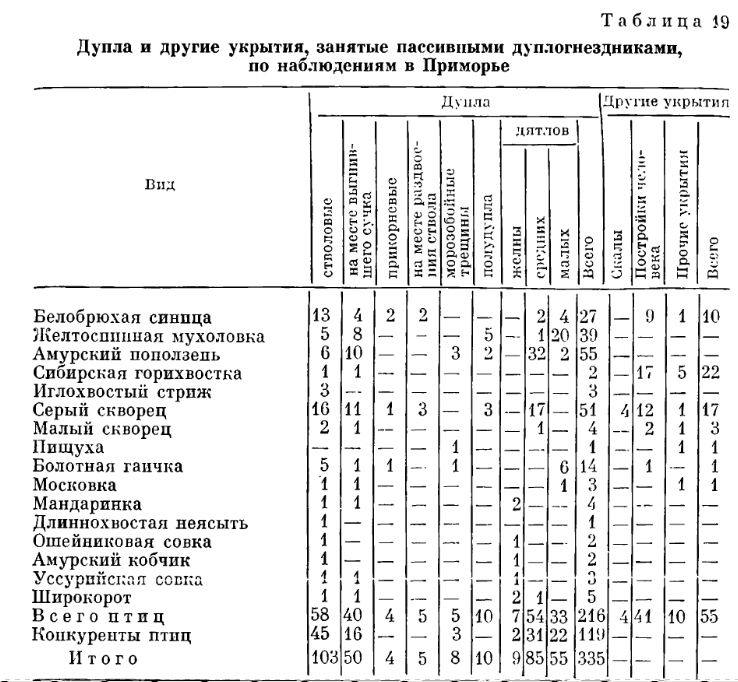
В. Заселение разных типов полостей (в том числе дупел природных и дятловых) вторичными дуплогнёздниками в Южном Приморье. Источник. Табл.19 в: В.М.Поливанов. Экология птиц-дуплогнёздников Приморья. М.: Наука, 1981. 173 с.
Рисунок 1. Гнездовые сети умеренных лесов Северной (А) и тропических (Б) лесов Южной Америки, а также юга Приморья (В).
Первая – пресечение резидентными особями потенциально возможных вспышек «вредителей». Важней всего здесь виды-специалисты (D.leucotos, P.tridactylus, D.medius и т.д.): только они используют кормовые методы, самые стереотипные по исполнению, многосоставные и наиболее сложные в осуществлении, долгие по времени, но и самые «уловистые» в добыче соответствующих жертв (почему и считаются специализированными, рис.2, табл. 2). Они обеспечивают наилучшее проникновение в соответствующий субстрат для их зондирования и схватывания [13, табл.2], включают в себя разные виды долбления и сбора с поверхности последнего. Специализированным видам дятлов структура субстрата (живая или мёртвая древесина, развитие корки с её сложной поверхностью, чечевички и механические повреждения и т. д.) важнее места кормления, в смысле конкретного типа леса и микростаций внутри кроны [особенно белоспинному и трёхпалому, 14-15]. Для генералистов (прежде всего D.major, D.villosus, D.pubescens), наоборот, местообитание важнее субстрата.
Действительно, трехпалые дятлы P.tridactylus, P.arcticus весьма эффективны в контроле численности короедов, белоспинный D.leucotos — заболонников, лубоедов и древоточцев, средний D.medius и малый пëстрый D.minor голых листогрызущих гусениц (первые два — круглый год, последние — в гнездовое время, когда ими же преимущественно выкармливают птенцов) [2, 12, 16] и т. д.

Рисунок 2. А. Гильдия дятлов в европейских лесах: разные специализации и размерные классы
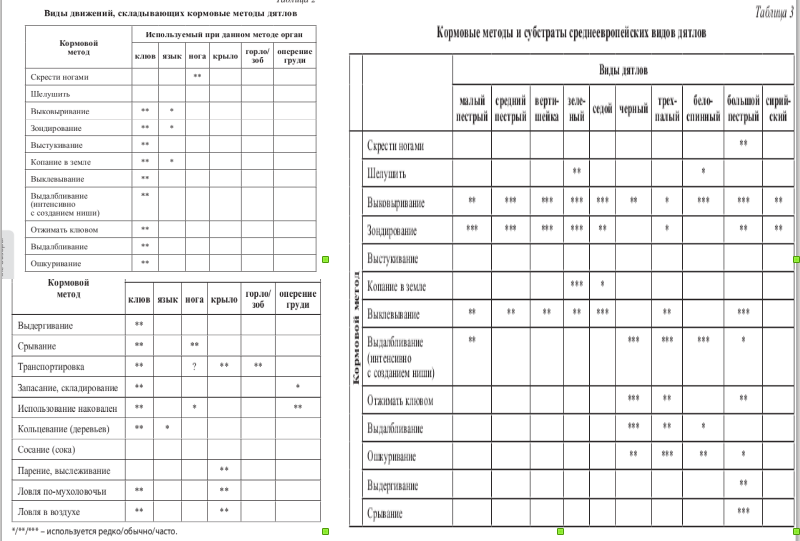
Б. Репертуар кормовых методов и их использование разными видами. Fig.2 в: Scherzinger W. Niche separation in european woodpeckers — reflecting natural developing of woodland// Intern. Woodpecker Symp. Nationalpark Bertechsgaden. 2002. Forschungsbericht 48. S.139-154; Табл.2-3 в: Фридман В.С., Суслов. Нетривиальные формы конкуренции:» шум» и» помехи» как средства борьбы за существование// Социально-экологические технологии. 2018. №4.
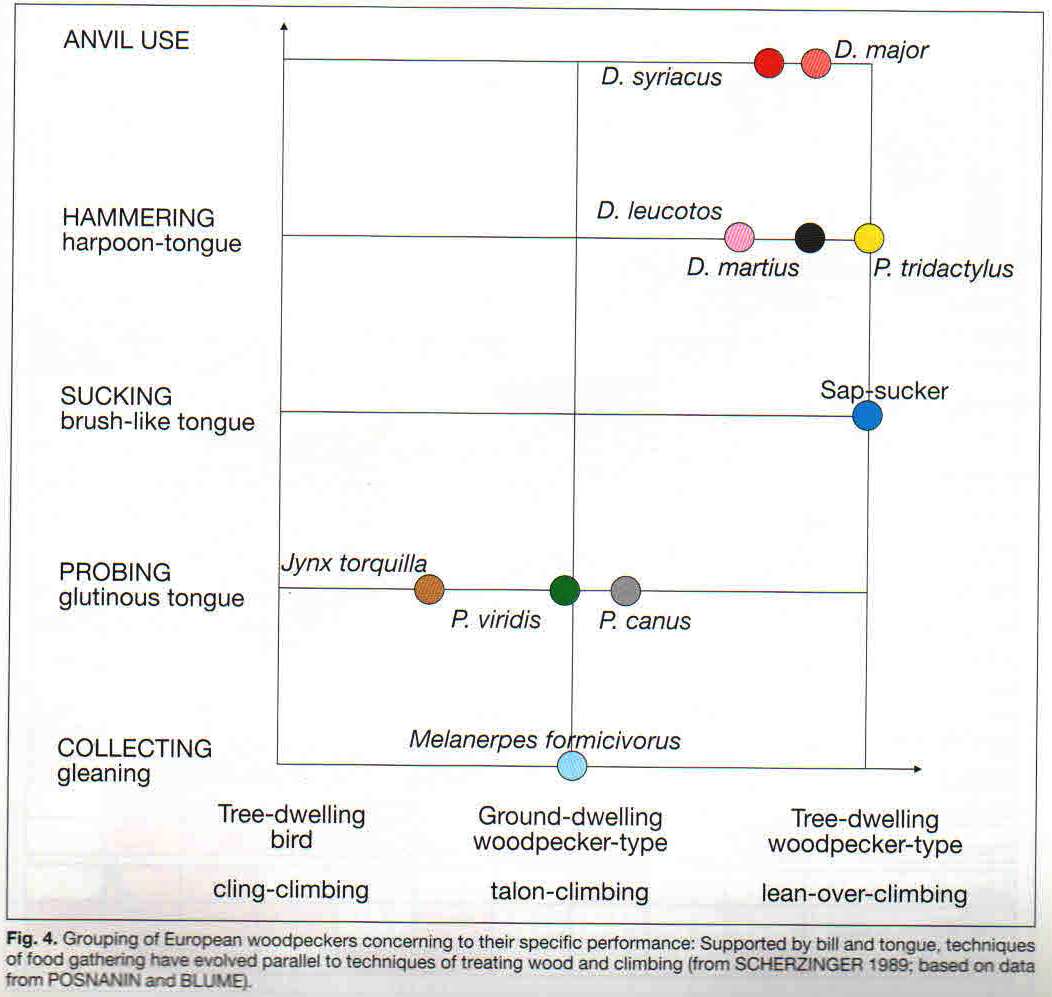
Рисунок 3. Экологические типы голарктических дятлов: специалисты-долбильщики на стволах, специалисты-собиратели на стволах, наземные специалисты-копатели (в том числе мирмекофаги), генералисты. Обозначения. Строки сверху вниз: использование кузниц, долбление с «загарпуниванием» жертвы крючьями ороговевшего языка, сосание древесного сока и щётковидный язык, зондирование субстрата и клейкий, особенно длинный язык. Столбцы слева направо: экологические группы дятлов: вертишейки (древесные, обхватывающе-лазающие виды), земляные дятлы (наземные виды, при лазаньи по стволу далеко отклоняющие тело, «висящие на когтях»), жёлны и настоящие дятлы (древесные, при лазаньи по стволу тело прижато к нему, ноги расставлены). Источник. Fig.4 в: Scherzinger W., op.cit.
Гильдии дятлов в лесных биомах: жизненные стратегии и специализация видов
Для разных видов дятлов и типов лесных биомов показана достаточность определённой численности вида-специалиста для исключения вспышек массового размножения его «типовых» жертв (горные ельники Швейцарии — трёхпалый дятел [17]). При меньшем уровне численности3, происходящую вспышку птицы не ликвидируют — места регулярного кормления владельцев участков «накрывают» не всё пространство массива, а они сами слишком привязаны к ним, слишком устойчиво следуют постоянным маршрутам каждодневных обследований, чтобы концентрироваться в очаге [13].
Так, белоспинный и трехпалый дятлы — индикаторы малонарушенных хвойных или лиственных лесов — встречаются на лесных участках площадью более 100 га с обилием сухостойной древесины не менее 20 м3/га. Это в пять раз меньше, чем в малонарушенных лесах, но в пять-десять больше, чем в эксплуатируемых. Такие леса, даже существенно изменённые и нарушенные (англ. seminatural forest, «полуприродные» леса) могут быть долговременными рефугиумами биоразнообразия [13]. Показана важность обилия мертвой древесины в местообитаниях среднего дятла, ее роль в выборе мест гнездования4 и как зондируемого субстрата наравне с коркой крупноразмерных дубов и грабов, вопреки прежним мнениям о кормлении Dendrocopos medius лишь на живых стволах [20].
Поэтому, кроме численности вида в данной местности, предотвращение вспышек прямо пропорционально жёсткости территориальных связей резидентной части популяции, их внутри- и межгодовому постоянству. Она максимальна у видов-специалистов, где многолетне-постоянные пары используют многолетне-постоянные же участки обитания: крупные (1-2 км2), с крайне стабильной конфигурацией как их самих, так и зоны использования внутри. Оба настолько постоянны, часто сохраняются при смене владельцев (случающейся раз в несколько лет, реже ежегодно), т. е. стабильны десятилетиями. См.данные по белоспинному и трёхпалому дятлам в сравнении с генералистом большим пёстрым [2, 13, 20].
Нетерриториальным особи тех же видов (они образуют «подвижный резерв» популяции), напротив, присущи дальние и быстрые перемещения, особенно самкам. Скажем, приток птиц из Карелии поддерживает уязвимую финскую популяцию D.leucotos, бывшую в 1990-2000-е на грани вымирания и восстанавливающуюся крайне медленно [21]. Перемещения трёхпалого дятла даже включают выходы за пределы лесной зоны, в лесные микрофрагменты (колки) лесостепи. В них могут участвовать и постоянные пары, начинающие размножение в местах остановок и т. д. [2]
У видов-генералистов (в Евразии большой пёстрый дятел и желна Dryocopus martius, в Неоарктике волосатый и пушистопёрый) участки резидентов много подвижней. Их общая конфигурация, привязка к местности, «пятна» функциональных зон внутри5 куда вариабельней а) в течение года, между разными стадиями годового цикла, б) в разные годы даже в случае, когда он используется несколько лет. Однако у данных видов он гораздо чаще переносится в другое место, или в другой массив при смене сезонов или на следующий год. Обычно это сопровождается распадом пар на осень-зиму, даже когда следующей весной они воссоединяются в прежнем составе. Иными словами, территориальные и брачные связи здесь много более «гибки», но их изменения «мельче» по масштабам захватываемых ими времени и пространству (бывает чаще, идёт на меньшие расстояния и пр.) [2].
Второй, лучше всего изученный, аспект «санитарной» роли дятлов в лесных биоценозах (иногда и не только [2]) — их способность концентрироваться в очаге размножения «вредителей», если вспышка уже произошла. Это делают особи «подвижного резерва» популяции: вместе со всей дендрофильной авифауной, прежде всего певчими птицами, они активно уничтожают корм, ставший вдруг массовым и легкодоступным6. Так мобилизуются все виды дятлов, специалисты и генералисты7: благодаря этому численность нетерриториальных особей, приходящих издалека, быстро растёт, как в ельниках Подмосковья, погибших от вспышек короеда-типографа, повторяющихся регулярно с 1998 г. в сухие жаркие лета. Концентрация дятлов, ликвидирующих очаг, идёт наиболее споро во внегнездовое время, особенно если вспышка совпала с недолгим сезоном весенних перемещений или, подольше, позднелетних, переходящих в осенние [22-23]. То же самое зафиксировано в смешанных или широколиственных лесах Центра Нечерноземья, бореальных и хвойно-широколиственных лесах Северной Америки8 и, видимо, представляет собой общее явление [2, 12, 24].
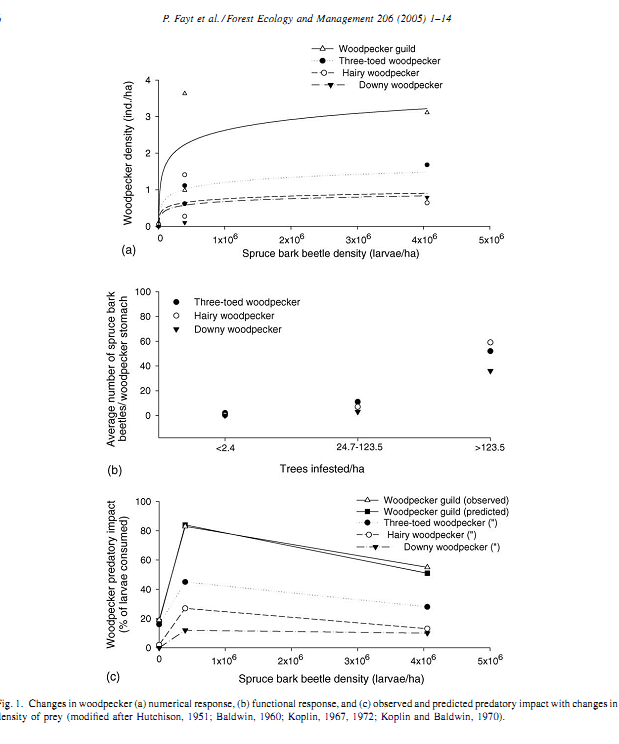
Рисунок 4. Концентрация 4-х видов дятлов на вспышках размножения короедов в бореальных лесах Северной Америки и их воздействие на «вредителя». а. Отклик изменением численности (особей/га) каждого из видов (трёхпалый дятел — чёрные кружки, волосатый дятел — белые кружки, пушистопёрый дятел — чёрные треугольники) и всей гильдии дятлов (белые треугольники) на рост локальной плотности короедов (личинок/га).
б. Функциональный отклик переключением на массовый корм. Показано среднее число личинок короедов в желудках разных видов дятлов (обозначения те же) в зависимости от плотности заражённых деревьев в данном массиве (экз./га). в. Интенсивность воздействия разных видов дятлов (обозначения те же) на короедов в очаге вспышки массового размножения (% уничтоженных личинок) в зависимости от плотности жертв, наблюдаемые значения в сравнении с предсказанием модели мобилизации разных видов дятлов (чёрные квадраты).
Примечание. Обратите внимание на большую эффективность действия специализированных видов во всех аспектах, в которых рассматривается их влияние на «вредителя». Источник. Fayt P., Machmer M.M., Steeger C. Regulation of spruce bark beetles by woodpeckers—a literature review// Forest Ecology and Management. 2005. V.206. №1-3. Р.1-14.
Разные виды дятлов кормятся здесь бок о бок, без следования присущим им биотопическим привязанностям и предпочитаемым кормовым методам, не говоря уж о соблюдении территориальных запретов (хорошо видимых у резидентов: поэтому разные виды дятлов часто сопровождают многовидовые синичьи стаи, но почти исключительно поодиночке, без кормления неподалёку особей разных видов). Специалисты и генералисты ведут себя здесь одинаково неспециализированно, главным образом собирают корм, высматривают и схватывают его или выковыривают, неглубоко зондируя разрушаюшиеся субстраты — древесину, почки, реже почву или опад на ней. Вероятность предотвращения вспышки массового размножения отдельных вредителей (короеды, листовертки, пяденицы) положительно связана с уровнем численности видов, специализирующихся на добыче каждого из них, а также — но заметно слабее — с общим уровнем численности всех дятловых птиц и разнообразием населения разных видов по Шеннону. Виды-специалисты также дают существенно больший отклик на вспышки массового размножения «своих» жертв, и интенсивнее истребляют их (трёхпалый и черноспинный дятлы в сравнении с волосатым и пушистопёрым) [11-12, 24, table 2, 4].
После ликвидации очага дятлы исчезают оттуда, не пробуя загнездиться весной в ближайших массивах9. Часто они не гнездятся даже тогда, когда очаг настолько велик, что служит источником корма в течении нескольких лет (там появляются лишь проходящие особи). Случаи гнездования в ближайших окрестностях, даже когда оно делается регулярным (у трёхпалого дятла вокруг короедников в Подмосковье в 2001-2003 гг. и позднее) идёт не за счёт их «осаживания», а прибытия резидентов из соседних массивов, где вид регулярно гнездится или начал гнездиться в последние годы [22-24].
В меньшинстве таких случаев отдельные особи из очага всё же «оседают» на гнездовании, и размножаются там на протяжении от одного до пяти сезонов, но потом всё равно исчезают. Виды-генералисты так делают значимо чаще, их гнездование продолжается дольше, у специалистов — значимо реже, если только они не гнездились рядом до вспышки, тогда резиденты захватывают её своими участками [данные по D.villosus, D.pubescens, P.tridactylus, P.arcticus прибывшим в короедники в хвойных лесах США, 24]). Не-концентрация резидентов и жёсткость территориальных связей последних объясняет два наблюдения, вроде бы констрастирующие между собой. По данным 15-летних исследований во внутренней части Британской Колумбии, в годы крупномасштабных вспышек массового размножения соснового лубоеда Dendroctonus ponderosae 6 местных видов дятлов (золотой, волосатый, пушистопёрый, американский трёхпалый дятлы P.t.dorsalis, хохлатая желна Dryocopus pileatus, красноголовый дятел-сосун Sphyrapicus varius nuchalis) увеличили численность, но не интенсифицировали размножение. В противоположность гаичкам с поползнями, значительно увеличившим продуктивность, у них не возрос ни размер кладки, ни число слётков на пару, несмотря на изобилие корма в этом году и в следующих [51].
Сходным образом население дятлов реагирует на выборочные рубки — в местах их проведения увеличивает численность (в сравнении с нетронутыми участками), но не продуктивность размножающихся там пар. Всем видам дятлов — специалистам и генералистам — присуще постоянство «вложений» пар в репродукцию, вопреки сильным колебаниям обилия и доступности ресурсов между годами и/или территориями [51]. Сравнение седого дятла с зелёным, большого пёстрого — с белоспинным или средним, малого — с трёхпалым и пр. чётко показывает, что размер кладки — и выводка — видов-генералистов значительно больше и сильней изменяем в зависимости от обстоятельств, чем у родственных им специалистов: в среднем 5-10 и 3-4 на успешное размножение [2].
В таком случае долговременные перераспределения птиц внутри популяций, в том числе мобилизация дятлов в ответ на локальные изменения древостоев «в лучшую сторону» (обилие корма, перестали удалять сухостой, деревья слабеют и сохнут от изменений климата и т. д.) достигается без интенсификации размножения, одним изменением выживаемости резидентов и/или перемещаемости нетерриториальных и не-местных особей из «резерва». Сухие и усыхающие деревья, а особенно подолбы на них здесь — не только физические объекты (возможные места кормления), но прежде всего — элементы сигнального поля, направляющие приток особей со стороны и делающие мобилизацию особенно быстрой.
В малонарушенных лесах обычные роли дятлов незначимы
Увы, накопление полевых данных показывает, что обе экосистемные «роли» дятлов значимы лишь во вторичных и производных лесах, часто происходящих из лесокультур или изменённых хозяйственным использованием10 (рекреационные леса; коммерческие леса11, фактически лесные плантации; леса, прошедшие через пашню и используемые на дрова, под сенокосы, огороды, в других лесных промыслах и пр.) в сторону однородности структуры и одновозрастности древостоев. Изучать биологию дятлов начали именно с них, что и канализировало впечатление исследователей.
Современное накопление данных по другим типам древостоев, включая сохранившиеся «обломки» малонарушенных лесов, развивающихся в спонтанном режиме12, и полуприродные леса (англ. seminatural forests), сохранившие известную долю структуры (а, значит, и облика) малонарушенных, показывают несущественность там обоих экологических функций дятлов. Если точней, там они выступают побочным следствием главной ценотической роли этих птиц – двусторонней «работы» по воспроизводству структуры лесного ценоза. Их средопреобразующая деятельность, зависящая от распределения и обилия листогрызущих и стволовых «вредителей» на разных субстратах, оставляющая «следы» в виде подолбов и других «погружений» (самые крупные/глубокие из которых — гнездовые и ночёвочные дупла в местах долговременного обитания) пропорционально интенсивности нападения «вредителей» на деревья одновременно
(*) сохраняет и дальше те парцеллы растительности (с типичной для них «архитектурой» крон, определёнными прорывами или окнами между ними), которые данным разнообразием видов дятлов могут быть защищены от данного нападения фитофагов (с учётом численности и кормовой специализации каждого вида), но;
(**) ускоряет усыхание (у мёртвых — слом и разложение) деревьев и групп деревьев, незащитимых от данного нападения фитофагов при данном разнообразии дятлов. Они же совместно с дереворазрушающими грибами обеспечивают «посмертие» деревьев-средообразователей, в каждый данный момент максимизируя образование мертвой древесины и поддерживая скорость её разложения на уровне, оптимальном для данного ценоза. Одновременно
(***) усиливается контраст «зеленеющих дальше» и ускоренно усыхающих, формирующих более или менее глубокие прорывы полога, куртин деревьев. Воспроизводясь активностью дятлов, он образует одну из эндогенных мозаик малонарушенного ценоза, «вкладывающуюся» в мозаики, созданные прочими эдификаторами [25-26], как одна матрёшка в другую, и располагаясь друг относительно друга как разные слои ГИС. Все вместе они формируют типичную картину пространственной структуры ценоза, где дятлы оказываются такими же эдификаторами, как зубры, туры или бобры, только действующими в меньшем масштабе времени и пространства.
Далее обосновывается данная точка зрения.

Рисунок 5. Принципиальные различия в распространении «пятен» лиственного леса (в таёжной зоне связанного с «окнами» нарушений в основном хвойном покрове) при естественной динамике лесного ландшафта (слева) и в эксплуатируемом лесу (справа). В природном ландшафте мелколиственный компонент встречается в виде примеси (отдельных деревьев или групп деревьев – точки), как преходящая стадия сукцессии (заштрихованные участки), как стабильные сети и полосы сырых лесов, протягивающихся вдоль водотоков. Из: [31]
Структурная сложность лесных ценозов: разные измерения
В малонарушенных лесах достигается максимум сложности структуры ценоза (англ. Naturalness, что можно перевести как сохранность природного облика). Она оценивается разнообразием разнотипных мозаик, слагающих лесной полог по вертикали (кроны разной архитектуры с более или менее глубокими прорывами» между ними) и по горизонтали (мозаика парцелл растительности разных ярусов, в том числе кустарникового и травянистого ярус, сопряжённая с таковой форм рельефа, типов почв и пр.). [13, 19, 25-27]. Последнее оценивается по нескольким измерениям, характеризующим эту структуру.
Во-первых, это разрежения древесного яруса во всем диапазоне их размеров и глубины: от полян и редин через «окна» от выпадения отживших своё деревьев первого яруса до разноразмерных «прорывов» полога, не доходящих до земли (от частичного усыхания, уничтожения части листы и пр.). Во-вторых, обилие (м2/га) и пространственное размещение разных видов мертвой древесины (сухостой поодиночке и группами, валеж и т. д.). В-третьих, разнообразие крон разных видов деревьев-средообразователей первого яруса: высот их расположения друг относительно друга вследствие смешанности и разновозрастности древостоя, сопряженное с разнообразием форм микро- и мезорельефа, положительных и отрицательных, сообразно отличиям их почвенно-влажностных условий [13, 19, 25-27]. Все три измеримых аспекта разнообразия достигают максимума в малонарушенных лесах и сокращаются пропорционально потере природного облика [25-26].
В-четвертых, здесь значимо разнообразие форм и размеров «вкраплений» мелколиственных видов деревьев в бореальных или широколиственных лесах, отражающих прошлое образование и зарастание «окон». В малонарушенных лесах их много, они разнородны, в том числе крупные и привязанные к соответствующим формам рельефа: «пятна» в бывших рединах и «окнах», «полосы» вдоль водотоков и заболачиваний, созданных активностью бобров Castor fiber. В коммерческих и других изменённых лесах они точечны и случайно разбросаны по массиву (если не формируют крупных однородных массивов березняков и осинников, также с низким структурным разнообразием). Противоположно распределяются более или менее восстановленные «пятна» нарушений эндогенного характера — ветровалы, пожарища, усыхание отдельных стволов от «вредителей», заболачивания с гибелью леса и пр. В лесах, развивающихся в спонтанном режиме, они минимальны по площади, точечны и случайно разбросаны по массиву, в нарушенных, наоборот, крупноразмерны и собраны в группы [13, 30-31].
Хозяйственная эксплуатация «накладывает» сеть внешних нарушений на эндогенную мозаичность, тем большую, чем сильнее нагрузка и разрушительней использование. Ее «пятна» крупнее, соединены друг с другом, обычно дорожно-тропиночной сетью. Всё это инсуляризует массив: что хуже всего, «островки» и «прожилки» антропогенных нарушений более или менее быстро «растут», ещё больше ветвясь и сливаясь между собой. Единый массив превращается в «архипелаг», в каждом «острове» это дробление с разъединением участков исходных сообществ нарушениями повторяется снова и снова [13, 54].
В пятых, в малонарушенных лесах максимум разнообразия полога «по вертикали»: разновысотность деревьев одного яруса, разнообразие диаметров стволов, измеренных на высоте груди; повышена доля крупноствольных и старых лиственных деревьев в зоне бореальных лесов, как и темнохвойных -в зоне широколиственных. Все пять «измерений» с разной стороны характеризуют структурную сложность ценоза, давая возможность количественной оценки «естественности» (или наоборот, производности и нарушенности) [19, 29-31].
Современные массивы такого рода (Беловежская пуща, разновозрастные дубравы «Калужских засек» и пр.) испытывали хозяйственную эксплуатацию в прошлом, утратили прежних крупнейших средопреобразователей, тура Bos primigenis и зубра Bison bonasus, и являют собой лишь некоторое приближение к доагрикультурным лесам, некогда простиравшихся по всей Восточноевропейской равнине, как они выглядят в реконструкциях [26, 28]. Структура лесного полога там была ещё разнообразнее и «ажурней» (больше разноразмерных «прорывов» и «окон» на единицу площади) за счёт ещё большей разновозрастности и разнопородности древостоя, а также включения нелесных участков. Внутри этой лесной полосы «от моря до моря» с севера на юг рос % площади, занятой «островами» лугово-степных участков, их средний размер и связность между, болотных и тундроподобных участков — в обратном направлении.
Они возникали за счёт средопреобразующей деятельности ключевых видов животных, прежде всего исчезнувших зубров и туров, ныне восстановленных и массовых — бобров, а также ряда других [25, табл.4]. Жизнедеятельность каждого из экосистемных инженеров табл.4 рождала своё «кружево» нарушений, каждое в своём масштабе пространства, со своей скоростью «зарастания» в ходе вторичных сукцессий. «Накладываясь» наподобие слоёв ГИС, включаясь друг в друга как матрёшки, «следы» их жизнедеятельности создают максимум эндогенной мозаичности ценоза [25, табл.1-3].
Ныне от него сохранились только «осколки» — однако достаточные для сложения в непротиворечивую реконструкцию и определения изменённости современных лесов сравнительно с этим образцом [26]. Количественно последняя оценивается как потеря «натуральности» лесного сообщества, разрушение и упрощение их исходно сложной структуры, с исчезновением мёртвой древесины, крупных и нестандартных деревьев, других элементов, важных для разнообразия местообитаний и устойчивости биогеоценоза. Максимум последнего достигается в коммерческих и рекреационных лесах Европы, с её интенсификацией и отдыха и хозяйствования: российские с приходом капитализма развиваются в ту же сторону13 [19, 29-30, 54-55].
В малонарушенных лесах дятлы как «производители» дупел не значимы
После «ранжирования» лесов по степени натуральности немедля оказывается, что при «фоновом» и даже среднем уровнях нарушений дятлы как «производители» дупел незначимы, потенциальным «жильцам» хватает естественных [4, 6, 9]. Это верно и для большинства «полуприродных» лесов. Они достаточно старые (спелые и приспевающие, реже средневозрастные), с вкраплениями мелколиственных пород, где сухостой, крупные и нестандартные деревья не удалены полностью, с промежуточными значениями разнообразия по 5 структурным характеристикам, обсуждённым выше, то есть с известной сохранностью компонентов, «оставшихся» от исходного биогеоценоза [32].
И в первых, и во вторых дятлы как «производители» дупел отнюдь не существенны, природных достаточно [4, 6, 9, 32, 33, рис.6-7]. Такие леса также достаточно захламлены, а часть «потребителей» в гнездовой сети может гнездиться и в полудуплах и других нишах/укрытиях, не являющихся дуплом [34]. Они могут быть значимы лишь в насаждениях, где дятловые дупла преобладают над естественными, т. е. в управляемых человеком лесах по сравнению с охраняемыми одного и того же ландшафта [52].
Таблица 1. «Предложение» дупел и факторы, на него влияющие, при рассмотрении в разном масштабе пространства (I — мета-анализ данных по лесам Европы и Северной Америки, II-IV — данные автора по разным местностям Восточной и Центральной Эстонии [53, fig.1].
Примечание. 2 и 3й столбцы — оппозиции, определяющие обилие дупел (больше-меньше), Эффект — влияние соответствующих факторов как переменных по данным дисперсионного анализа, М — много- и U-однофакторного. Везде представлены средние значения, * — — их разница статистически значима.
| Масштаб | Больше дупел в: | Меньше дупел в: | Эффект | Регион |
| Континент | Северной Америке | Европе | Дупел: 17.7 га –1 vs. 5.6 га –1 (M*) | I |
| Влажном климате | Умеренном и сухом климате | Дупел: 19.6 га–1 vs. 5.9 га–1 (M*) | I | |
| Ландшафт/местность | Спелые леса | Средневозрастные и молодые леса | Дуплистые деревья: 1.7 га–1 vs. 0.5 га–1 (U*) | II |
| Лиственные и смешанные леса | Хвойные леса | Дуплистые деревья: 0.7 га–1 vs. 0.5 га–1 (U*) | II | |
| Природные леса | Управляемые леса | Дуплистые деревья: 1.3 га–1 vs. 0.8 га–1 (M) | II | |
| Поблизости от других дупел | Дуплистые деревья не распределены равномерно по массиву, а образуют скопления: 56% из них расположены на 10% территории (U) | II | ||
| Насаждения | В радиусе 10 м вокруг дупла плотность дятловых и природных выше средней для массива в 12 и 4 раза соответственно (U*). | IV | ||
| Природные древостои | Управляемые древостои | Дупла: 18.3 га–1 vs. 5.4 га–1 (M*) | ||
| Осинники | Березняки | Дупла: 7.5 га–1 vs. 1.6 га–1 (M*) | ||
| Деревья | Около лесной опушки | Во внутренней части леса | Встречаемость дупел в 3 раза выше в 50метровой краевой зоне массива по сравнению с центральной частью (M*) | |
| Осина и чёрная ольха | Берёза | Встречаемость дупел в 4 и 2 раза выше средней по массиву vs в 2 раза ниже средней (M*) | ||
| Сухостой | Живое дерево | 8кратные различия по встречаемости дупел в первых сравнительно со вторыми (M*) | ||
| Крупные деревья | Тонкие деревья | Обилие дупел в 4 раза больше в стволах с DBH>30 см (M*) | ||
| Осины, дубы, буки и сосны | Ели и берёзы | Если деревья данных родов присутствуют в составе древостоя, в них оказывается 100-43% дупел vs 0% | ||
Источник. Remm J. Tree-cavities in forests: density, characteristics and occupancy by animals. Tartu: Tartu University Press, 2008. P.19.
Больше того, для долбления дупел дятлы предпочитают именно те субстраты, где вероятней всего пойдёт и естественное образование дупел, пусть много медленней:
а) стволы и участки ствола с наиболее мягкой древесиной (сравнительно с прочими видами деревьев или с другими участками того же ствола/сука) [35];
б) даже твёрдодревесинные виды деревьев, если их ствол и особенно сердцевина размягчены дереворазрушаюшими грибами или бактериальными гнилями, распространившимися через трещины, морозобоины, сломанные или выгнившие сучки и пр. «ворота» инфекции [36]. От а)-б) дятлы сильно зависят: выбирают ствол, а на нём — место строительства гнезда, строго ориентируясь на оба критерия. Зависимость настолько сильна, что птицы бросают почти завершённое дупло, наткнувшись на внутренний сук, свилеватость, другой участок древесины, не утратившей «здоровой» жёсткости [35-37].
Дятлы не просто предпочитают деревья, где древесина изменена теми же процессами, что обуславливают образование естественных дупел. По той же причине образовавшиеся пары ищут место для гнезда, позже и строят его именно там, где вероятность найти природное дупло (или старое собственное) также наибольшая: в местах наибольшей концентрации сухих или усыхающих деревьев, частично повреждённых или заселённых грибами,если сравнивать со случайно выбранными участками того же массива [35-37]. Первое важно вторичным дуплогнёздникам, второе — самим резидентам.
Так, в рекреационном Венском лесу большинство дупел выдалбливалось дятлами, естественные составили ~16%. Они концентрировались в участках возрастом 170 лет. L.M.Sachlerner [57] исследовал выбор там мест гнездования мухоловкой-белошейкой Ficedula albicollis. Участки вокруг её гнёзд отличались от наугад выбранных внутри массива: мёртвой древесины было больше, как сухостоя, так и валежа, выше концентрация дупел на 1 дерево, а их высота ниже. Почему относительное предпочтение естественных дупел белошейками было выше, а дятловых — ниже, чем «предложение» тех и других в биотопе [57].
Иными словами, в градиенте местообитаний разного качества дятлы с большей вероятностью разместят гнездо там, где вторичные дуплогнёздники, скорее всего, найдут и естественное дупло, а не там, где последние в дефиците. Выбор мест гнездования у них неслучаен в том смысле, что снижает зависимость потенциальных «жильцов» от их дупел. Последняя максимальна лишь у крупнейших из них — клинтуха Columba oenas, серой неясыти Strix aluco14, мохноногого сыча Aegolius funereus, гоголя Bucephala clangula, мандаринки Aix galericulata, сильно зависящих от желны (а их близкие родственники в Северной Америке — от хохлатой желны Dryocopus pileatus)15. Из 42-х жупел желны, прослеженных в 1989-1993 гг. в пригородных лесах Липецкой обл., с рекреационной нагрузкой от значительной до умеренной, 6 (14,3%) заселяли сами дятлы, 17 (40,5%) - лесные куницы, 5 (11,9%) - клинтухи, 3 (7,1%) - белки, 3 (7,1%) - серые неясыти и 8 (19,1%) пустовали. В лесопарках заселённость куницами, опасными для самих дятлов, была ещё выше (до 77,8%) [58].
В случае многолетнего обитания (то есть при наибольшей возможности увеличения «фонда дупел» для других обитателей леса) разные виды дятлов «сачкуют», учащают повторное использование дупел (и многодупельных деревьев) в силу его выгодности. Оно экономит время и силы: гнездование в старых дуплах начинается раньше и при прочих равных более продуктивно, лишь бы их было много и не очень разрушенных, в том числе прошлыми обитателями [5, 7].
Дятлы делают слишком «долгоживущие» дупла: выдалбливают их в слишком толстых стволах, тратя на это силы и время, при том что могли бы и меньше. Особенно виды — генералисты: большой пёстрый, обыкновенная и хохлатая желна, волосатый дятел. Это снижает зависимость от них вторичных дуплогнёздников: последовательно принимая разных «жильцов», дятловые дупла сохраняются первые несколько лет, в ряде случаев 10-20 лет [4].
Однако, чем лучше условия для выдалбливания, тем меньше «живёт» дупло, выдолбленное в подобном стволе: в сухостое меньше, чем в усыхающих, а там — чем в живых стволах, в дубово-грабовом лесу или прибрежном лесу меньше, чем в ельнике на плакоре и пр. [4]. Возможно, поэтому естественные дупла долговечней. В Беловежской пуще дятловые дупла сохраняются максимум 31 год, естественные — 36 лет, но некоторые разрушаются ещё до начала следующего сезона гнездования. Медианное значение сохраняемости естественных дупел — 12 лет, дятловых — 7 (табл.2). Следовательно, первые «живут» значимо дольше: на деле эти различия ещё сильней, т. к. возраст естественных дупел сильно недооценивается, отсчитываясь от первого известного использования [9].
«Срок жизни» дупел варьирует в зависимости от субстрата и биотопа: дятловые дупла в хвойных сохраняются дольше, чем в лиственных (10 и 6-7 лет), тогда как естественные — наоборот (4,4 года — 10-13 лет). Напротив, в Британской Колумбии оба типа дупел в осинах сохраняются до 21 года. Сохраняемость выше у дупел в живых деревьях, нежели в мёртвых, и ниже у сделанных «слабым выдалбливателем» сравнительно с «сильным». Разница в сохраняемости связана с распространённостью гниения в субстрате, где выдалбливается дупло: чем сильней им размягчена древесина, тем недолговечней дупло [9].
![Таблица 2. Медианная сохраняемость естественных дупел в разных регионах, лет. Обозначения: строчки сверху вниз: Беловежская пуща (польская часть), Эстония, Британская Колумбия, Иллинойс, США; Колорадо, США. Столбцы, слева направо: длительность наблюдений, лет; медианная сохраняемость, лет; источник. А - Экстраполяция по данным, обозреваемым в соотв.работе. Источник. Табл.4.2 в [9]. В малонарушенных лесах деревья, сильно поражённые грибами — лучшие индикаторы](http://www.socialcompas.com/wp-content/uploads/2021/11/Snimok-ekrana-ot-2021-10-26-19-17-46.png)
Таблица 2. Медианная сохраняемость естественных дупел в разных регионах, лет.
Обозначения: строчки сверху вниз: Беловежская пуща (польская часть), Эстония, Британская Колумбия, Иллинойс, США; Колорадо, США. Столбцы, слева направо: длительность наблюдений, лет; медианная сохраняемость, лет; А — Экстраполяция по данным, обозреваемым в соотв.работе. Источник. Табл.4.2 в [9].
В малонарушенных лесах деревья, сильно поражённые грибами — лучшие индикаторы дятловых дупел [8, 11, 36]. Видимо, по той же причине, живые деревья с дуплами дятлов особенно привлекают короедов (живущих в симбиозе с грибами). Заражение ими обоими приближает гибель дерева, сокращает «срок службы» дупла, что бывает существенно для популяции. В Техасе поражение южным сосновым короедом Dendroctonus frontalis сосен ладанной Pinus taeda и короткохвойной P.echinata с гнёздами охраняемого кокардового дятла D.borealis – основная причина их гибели. Общее число заражённых и погибших деревьев с дуплами варьирует по годам пропорционально численности “вредителя”, предпочитающего именно их, независимо от того, выдолбили дупло сами птицы, или оно искусственное (вставлен сегмент ствола с дуплом). За 5 лет так погибли 20 сосен с первыми (25,6%) и 19 со вторыми (18,8%, различия не значимы); 25 из 62 сосен с дуплами, погибших от короеда в 1984-1996 гг., были гнездовыми в предыдущий сезон [56]. Наконец, долбление дятлов сокращает «срок жизни» дупел, когда приходится на гнездовые деревья: сделанные подолбы ослабляют прочность ствола. Они же увеличивают риск слома дерева «по дуплу» (главнейшая причина гибели дупел) и способствуют гниению древесины, особенно там, где она и так истончена [4, table 1].
Краснохохлый или кокардовый дятел

Иными словами, «производство» дятловых дупел везде детерминировано процессами, «рождающими» и природные полости: усыханием деревьев с возрастом или от нападений «вредителей», гниением древесины под действием дереворазрушающих грибов или бактерий, распространяющемуся по физическим повреждениям — трещинам, морозобоинам, кормовым подолбам дятлов и пр. Первое сильно облегчается вторым и крайне затруднено ему вопреки. Поэтому в большинстве древостоев «производство» естественных дупел «опережает» таковое дятловых: тем больше, чем более их возраст, местный климат, спонтанный режим развития биоценоза и прочие обстоятельства способствуют фаутности (если только самые подходящие деревья не удалены при экоопасном ведении лесного хозяйства и/или не уничтожены отдыхающими [4, 57]). Зависимость гнездостроения дятлов от наиболее мягкой и/или разрушенной гниением древесины сокращает «срок жизни» дупел именно там, где их больше всего — в приречных местообитаниях, в массивах с обилием мёртвых деревьев и пр. В Эстонии это 36 лет у живых деревьев на плакоре и лишь 6 — у сухостоя в пойменных биотопах. Дополнительно, эффективные «контролёры» массового размножения филло- и ксилофагов — малый, трёхпалый, белоспинный дятлы — особенно предпочитают мёртвую древесину. Поэтому их дупла самые «короткоживущие», и полезны другим видам ещё короче — первые год-два после постройки [4, 7, 38-39, 52].
Всё вышеизложенное определяет, что «предложение» дятловых дупел — не независимая величина, но функция «предложения» природных дупел: зависимость между ними описывается кривой с насыщением. Дятловые дупла преобладают в лесах, бедных дуплами вообще (управляемых и рекреационных), с ростом «фонда» природных дупел их доля экспоненциально падает. В малонарушенных лесах Европы и Северной Америки большинство вторичных дуплогнёздников используют природные дупла [9].
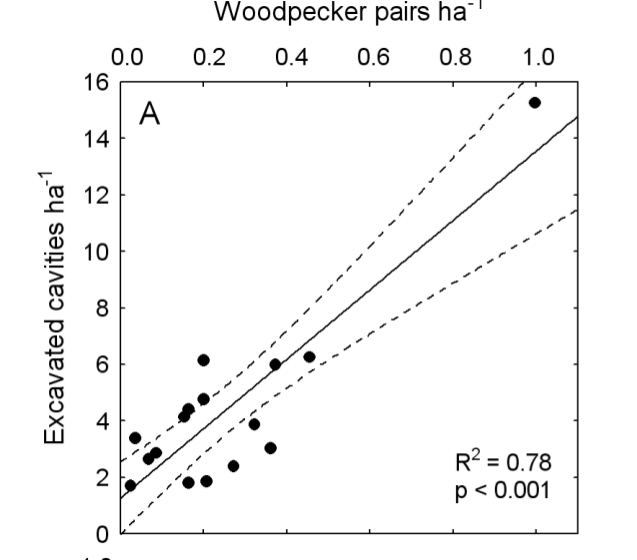
«Производство» дятлами дупел линейно зависит от плотности их населения, она же суть функция плотности природных дупел (фактически обилия мёртвой древесины), что также описывается кривой с насыщением. Поэтому плотность дятлов и их дупел имеет предел в 10-20 пар/га, но плотности природных дупел предела нет [6, 52]. Это верно как для объединённых данных по всем континентам (рис.6), так и для отдельных локаций (Эстония, [52] и рис.7).
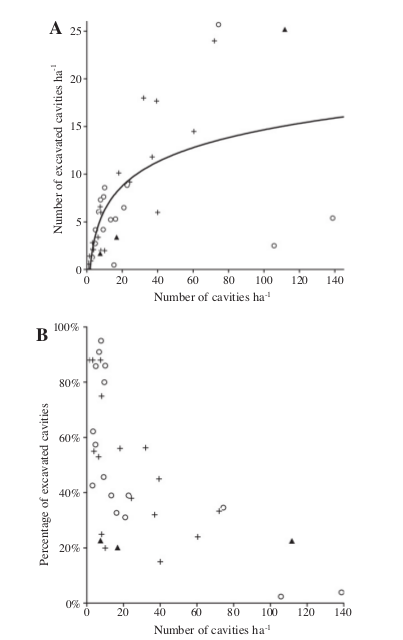 Рисунок 6. Связь между выдалбливаемыми (в основном дятлами) и природными дуплами для большинства лесных биомов. Ось Х — общее обилие дупел, га-1, ось У — число дятловых дупел (верхний) и их доля в % от общего «фонда») нижний рисунок. Обозначения: белые кружки — Северная Америка, + — Палеарктика, треугольники — неотропическая область. Примечание. Регрессия основана на лог-трансформации данных об общем обилии дупел, R21,27.0 = 0,51; P<0,001- Из: [6, fig.2].
Рисунок 6. Связь между выдалбливаемыми (в основном дятлами) и природными дуплами для большинства лесных биомов. Ось Х — общее обилие дупел, га-1, ось У — число дятловых дупел (верхний) и их доля в % от общего «фонда») нижний рисунок. Обозначения: белые кружки — Северная Америка, + — Палеарктика, треугольники — неотропическая область. Примечание. Регрессия основана на лог-трансформации данных об общем обилии дупел, R21,27.0 = 0,51; P<0,001- Из: [6, fig.2].
 А.
А.
Б.
Рисунок 7. Плотность дятловых (Б) и природных (А) дупел в зависимости от плотности гнездования дятлов на 16 приречных участках (абсцисса, пар/га), аппроксимация линейной регрессией, показан 95 % доверительный интервал. Значимость связи сохранилось и после исключения из анализа точки с наибольшей плотностью дятлов и дупел. Из: [52].
По всему миру плотность дятловых дупел значимо выше в климатических и ландшафтных условиях, способствующих образованию гнилей. Это повышенные температура и влажность, а также приречные и особенно пойменные местообитания [6, 8]. Леса Австралийской области, где дятлы (и прочие «изготовители») отсутствуют, наиболее изобильны последними, даже сравнительно с Неотропической областью, где больше всего видов дятлов на единицу площади. Велико здесь и разнообразие вторичных дуплогнёздников [52,2 га-1 vs 13,5 га-1 при сравнимом 95% доверительном интервале вокруг средней, 6, fig.3]. Долбление дупел критически важно лишь в некоторых лесах: достаточно молодых, с однородными древостоями, где «захламленность» устранена санитарными рубками и рубками ухода, исключивших использование естественных дупел [4, 9].
Наконец, вторичным дуплогнёздникам важнее разнообразие, чем обилие дупел, дятловые «подходят» не всегда и не всем. Изменение доли занятых ими дупел в зависимости от их численности описывается кривой с насыщением, причём на достаточно низком уровне: «лучших» дупел немного, за них интенсивно конкурируют, но большинство дупел пустует. Поэтому вторичные дуплогнёздники активно занимают искусственные гнездовья, иногда это делают даже дятлы [2, 8, табл.21, 5]. Отнюдь не все «популярные» дупел — дятловые, и даже не большинство: и общая численность, и разнообразие (по индексу Шеннона) гильдии птиц-вторичных дуплогнёздников линейно зависят не от обилия, но разнообразия дупел — по типу конструкции, форме дупла, виду дерева и месту расположения на нём и прочим характеристикам16. Дятловые дупла вполне однотипны и дают лишь один из нескольких занимаемых птицами вариантов [9, 52].
Возможно, поэтому в пойменных лесах Эстонии дятлы выдалбливают большую часть дупел, потенциально пригодных для воробьиных птиц, составляющих большинство вторичных дуплогнёздников. Однако те их в основном избегают, несмотря на обильное «предложение»; доля дятловых дупел в последнем фактически не влияет на численность и разнообразие видов вторичных дуплогнездников тех же лесов, при небольшой значимости разнообразия дупел [52, fig.5].
Из множества видов этой группы только у поползня Sitta europaea численность положительно коррелирует с обилием дятловых дупел [52]. Свою роль здесь играет хищничество некоторых больших пёстрых дятлов. Они раздалбливают дупла, известные им на участке, поедая птенцов занявших их видов (в том числе и других дятлов помельче). Заняв выдолбленные им дупла, певчие птицы фактически «подставляются» под эту угрозу. И наоборот, поползень выделяется обмазыванием летка глиной, что делает гнездо много менее доступным для хищников [8-9].


 Рисунок 8. Насколько разнородны процессы, ведущие к появлению и преобразованию дупел! Обозначения. (a) только что завершённое дупло красноголового дятла-сосуна Sphyrapicus nuchalis в живом тополе осинолистном, чуть повыше — инициальное дупло17. (b) щелевые дупла, созданные выгниванием, и заселяемые мелкими видами, в данном случае лазоревкой Parus caeruleus. (с) дупла на месте выгнивших суков: ветвь отрывается дальше или ближе к стволу, обнажается сердцевина, по ней распространяется гниль, в т.ч. вовнутрь узла, поэтому такие дупла бывают и в ветвях, и в самом стволе. (d) морозобоина, используемая обыкновенной пищухой. (e) дупла в живых деревьях зарастают, птицы могут препятствовать этому, удаляя каллус и обновляя внутренности дупла, как в этом старом дупле золотого дятла Colaptes auratus. Вверху слева — его инициальное дупло. (f) пока дупло, созданное выгниванием сердцевины, окружено здоровой древесиной, оно недоступно для разрушения, но дятлы, особенно большой пёстрый, могут продолбить туда новый вход, и использовать уже готовую полость. Все отверстия даны не в масштабе.
Рисунок 8. Насколько разнородны процессы, ведущие к появлению и преобразованию дупел! Обозначения. (a) только что завершённое дупло красноголового дятла-сосуна Sphyrapicus nuchalis в живом тополе осинолистном, чуть повыше — инициальное дупло17. (b) щелевые дупла, созданные выгниванием, и заселяемые мелкими видами, в данном случае лазоревкой Parus caeruleus. (с) дупла на месте выгнивших суков: ветвь отрывается дальше или ближе к стволу, обнажается сердцевина, по ней распространяется гниль, в т.ч. вовнутрь узла, поэтому такие дупла бывают и в ветвях, и в самом стволе. (d) морозобоина, используемая обыкновенной пищухой. (e) дупла в живых деревьях зарастают, птицы могут препятствовать этому, удаляя каллус и обновляя внутренности дупла, как в этом старом дупле золотого дятла Colaptes auratus. Вверху слева — его инициальное дупло. (f) пока дупло, созданное выгниванием сердцевины, окружено здоровой древесиной, оно недоступно для разрушения, но дятлы, особенно большой пёстрый, могут продолбить туда новый вход, и использовать уже готовую полость. Все отверстия даны не в масштабе.
Источник. Fig.4.2 в [9].
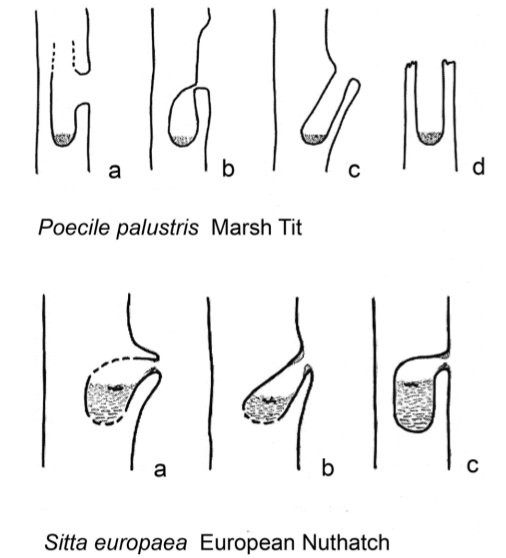 Рисунок 9. Продольные разрезы разных дупел (схематизированные, но с сохранением пропорций), занятых болотной гаичкой (вверху) и поползнем (внизу) в Беловежской пуще. У гаички это а) стволовое дупло в гнилой заболони, щелевидный леток, вход в вертикальной плоскости; б) то же самое, но вход в горизонтальной плоскости; в) дупло в узле, на месте выгнившего сука, д) «дымоход», со входом сверху по диагонали. У поползня а) дупло в узле на месте выгнившего сука, погружённого в конуссообразный нарост на стволе, б) то же, вход сверху по диагонали, леток уменьшен обмазыванием глиной, в) дятловое дупло, леток уменьшен обмазыванием глиной. Источник. Fig.4.3 в [9].
Рисунок 9. Продольные разрезы разных дупел (схематизированные, но с сохранением пропорций), занятых болотной гаичкой (вверху) и поползнем (внизу) в Беловежской пуще. У гаички это а) стволовое дупло в гнилой заболони, щелевидный леток, вход в вертикальной плоскости; б) то же самое, но вход в горизонтальной плоскости; в) дупло в узле, на месте выгнившего сука, д) «дымоход», со входом сверху по диагонали. У поползня а) дупло в узле на месте выгнившего сука, погружённого в конуссообразный нарост на стволе, б) то же, вход сверху по диагонали, леток уменьшен обмазыванием глиной, в) дятловое дупло, леток уменьшен обмазыванием глиной. Источник. Fig.4.3 в [9].
Дятлы как «санитары» значимы только в нарушенных лесах
В лесах, развивающихся в спонтанном режиме, дятлы в данной роли попросту не нужны: филло- и ксилофаги практически не дают вспышек из-за множества регуляторов. Хотя именно здесь в фаунистическом комплексе дятлов доминируют виды-специалисты, численность генералистов на минимуме, население — неустойчиво между годами (и часто меняет местообитания) [2, 13, 31]. Контроль насекомых, угрожающих древесному ярусу, важнее всего в seminatural forests: сохраняя природный облик, они нарушены достаточно для повторения вспышек [40].

Белоголовые дятлы: ухаживание в паре
С ростом антропогенной трансформации древостоев (урбанизация, лесоэксплуатация, рекреационная нагрузка) их пресечение делается всё важнее, но гильдия дятлов всё меньше способна это осуществить. Уменьшение натуральности лесных биомов меняет фаунистический комплекс дятлов — обедняет его и даёт перевес видам-генералистам. В пределе остаются только D.major с D.martius, в Америке — D.villosus с D.pubescens. Видовое богатство падает в ряду нарушенности и/или фрагментированности массивов за счёт выпадения видов-специалистов18, при сокращении уровня численности оставшихся [2, 13]. В этих условиях резидентная часть популяций оставшихся видов не может вспышку предотвратить, «подвижный резерв» популяций может концентрироваться в очаге начавшейся вспышки, но не может её погасить.

Американский трёхпалый дятел
Средообразующая роль дятлов: воспроизводство мозаики живых vs мёртвых деревьев
Ограничившись ролями 1) и 2), мы упускаем из виду главное — средопреобразующую деятельность дятлов. Она участвует в воспроизводстве лесного ценоза с присущей ему эндогенной мозаикой, причём двусторонне. В парцеллах, где древостой остаточно жизнеспособен при данной интенсивности нападений разнотипных «вредителей», кормодобывание дятлов — долбление, собирание и зондирование на разных видов деревьев, на разных субстратах и в разных микростациях внутри кроны [13, табл.2] минимизирует риск гибели дерева и появления прорывов полога в ближайшем будущем из-за потери большей части листвы и/или усыхания стволов. Если нет — то же самое действие тех же видов дятлов, сообитающих на данной территории, ослабляет деревья, дополнительно «открывая ворота» бактериальным и грибным инфекциям. Они ускоренно гибнут, или частично усыхают/теряют листву, ускоряя образование «окон» и разложение отмирающей при том древесины, но одновременно пресекается распространение этого на ближайшие участки.
Имущему воздастся, у неимущего отнимется: одной и той же активностью фаунистический комплекс дятлов, обитающих на данной территории, «укрепляет» деревья, выдерживающие давление фитофагов (увеличивает жизненность, снижает интенсивность нападений и пр.), но ускоряет гибель более слабых (у погибших — разложение древесины). Что, естественно, увеличивает «крупномасштабную» мозаичность ценоза во всех измерениях пропорционально биоразнообразию дятловых птиц каждого лесного участка. Благодаря этому дятлы заслуживают включения в список экосистемных инженеров [26, табл.4] наряду с другими животными.
Схожи последствия у долбления дупел: в отношении перспектив данного дерева это «самый крупный и глубокий подолб». Оно повышает риск слома ствола по дуплу (главная причина утраты старых дупел), особенно в случае сухостоя. Как при долблении с целью кормодобывания [41], здесь дятлы действуют совместно с дереворазрушающими грибами. Их клюв, проникая в субстрат, выступает вектором переноса спор вглубь ствола [42]. И наоборот, по другому преломлению света дятлы различают участки наиболее поражённые гнилью, и начинают кормление — или изготовление дупел — именно с них [43].

Темноспинный трёхпалый дятел
Так средопреобразующая деятельность дятлов «отделяет злаки от плевел»: деревья и их куртины, способные жить дальше при данной интенсивности нападений наличных «вредителей», от усыхающих, превращающихся в мёртвую древесину, с ростом жизнеспособности первых и интенсификацией разложения второй. Они или защитят дерево от «вредителей», если его жизненность выше порога (снижающегося, если интенсивность нападений растёт, или при учащении внешних нарушений вроде пожаров, механических повреждений, экстремальной погоды, а ныне — и ослабляющих дерево долговременных изменений климата [63]). Или те же воздействия приведут к противоположному результату: ускорят усыхание, у погибших — падение, у упавших — начало включения в кругооборот вещества и энергии [64].
Оба аспекта (особенно второй) зависимы от «покрытия» подолбами дятлов, их плотности и глубины. Это было показано экспериментально, именно из-за сопряжения с грибами [41]. Дятловое проникновение вглубь древесины, кормовое и гнездостроительное, вместе со сбором пищи с её поверхности, создают один из «узоров» эндогенной мозаики ценоза, поддерживают контраст между полноценным пологом крон и прорывами или даже «окнами» в месте удавшихся нападений «вредителей». Последние тем более точечны и случайно распределены по массиву, чем натуральнее лес и/или выше разнообразие видов дятлов, совместно его населяющих. И наоборот, чем более лес изменён и/или нарушен (либо чем меньше разнообразие дятлов, чем сильнее оно изменено по сравнению с потенциальным максимумом), тем они концентрированней и больше по площади, т.е. превращаются в очаг вспышки массового размножения.

Дятел Наттела, самец. Калифорния
Следовательно, на одной и той же территории и одновременно наличное разнообразие дятловых птиц выступает в двух взаимно-дополнительных экосистемных ролях. На нежизнеспособных участках они «пополняют запасы» мёртвой древесины, критически важные для лесного ценоза; «обрабатывая» её и перенося грибную инфекцию, они ускоряют «посмертие дерева». На жизнеспособных, наоборот, они сохраняют деревья разных ярусов, чем гарантируют сохранение сомкнутого полога и жизнеспособность листвы также и в будущем. Одновременно долбление ускоряет разложение мёртвой древесины, образовавшейся здесь ранее, в том числе валежа [41].
Следовательно, одной и той же активностью дятлы формируют разрежения полога или, наоборот, сохраняют ему сомкнутость дальше. Эта мозаика «санации» vs отмирания лесной растительности наряду с созданной прочими средопреобразователями на равных участвует в устойчивом воспроизводстве лесного ценоза, с присущей ему парцеллярной и ярусно-оконной структурой растительности, формами микро-, мезорельефа и пр. Особенно малонарушенных лесов, где она сложней и разнообразней всего [19, 30, 28], при постояннном балансе между а) образованием всех типов «окон» и «прорывов» полога кроны и б) их зарастанием в ходе восстановительных сукцессий, с равномерным распределением «пятен» а) и б) по площади массива.
Понятно значение для того и другого именно разнообразия дятлов, а не отдельных видов данной группы: чем больше последних выпало из орнитокомплекса от антропогенного изменения древостоя и прочих причин, тем больше вероятность образования каких-то «прорывов полога» без компенсации. И наоборот, чем больше видов обитает совместно, «обрабатывая» одновременно разными способами разные микростации, тем сложней и мелкомасштабней возникающая архитектура полога, а образующие её «прорывы» и окна меньше по площади и равномерней распределены по массиву.
Это лучше всего объясняет роль дятлов как индикаторов «натуральности» лесного биогеоценоза [44]. В меру роста структурной сложности лесных биомов растут продуктивность и фитомасса данных сообществ, давая возможность всё большему числу видов (и большему разнообразию экологических форм) дятлов обитать на данной территории. Поэтому их разнообразие увеличивается от тайги к влажным тропическим лесам — и наоборот, сокращается пропорционально фрагментации и нарушенности внутри массива [45-46]. Оно же в разных типах лесных сообществ выступает предиктором разнообразия дендрофильной фауны птиц (рис.10), здоровья леса, запасов его древесины [47-49], участки видов-специалистов, например, белоспинного — разнообразия беспозвоночных, связанных с мёртвой древесиной [50].
![Рисунок 10. Дятлы как предикторы разнообразия лесных птиц. а. В лесах умеренной зоны (сверху вниз — северо-восток Польши (n=33 площадки), восток Польши (т=33), Карпаты (n = 34), данные картирования для атласа птиц Польши в целом, чёрточки — ошибка средней (n=2317). Обозначения. Ось Х — число видов дятлов, ось У — число других лесных видов птиц. Источник. [47], Fig.3-4](http://www.socialcompas.com/wp-content/uploads/2021/11/Snimok-ekrana-ot-2021-11-04-12-07-02.png)
Рисунок 10. Дятлы как предикторы разнообразия лесных птиц.
а. В лесах умеренной зоны (сверху вниз — северо-восток Польши (n=33 площадки), восток Польши (т=33), Карпаты (n = 34), данные картирования для атласа птиц Польши в целом, чёрточки — ошибка средней (n=2317). Обозначения. Ось Х — число видов дятлов, ось У — число других лесных видов птиц. Источник. [47], Fig.3-4
![б. В тропических лесах (12 мест наблюдений в разных локациях. Строки сверху вниз: Неотропическая обл., Афротропическая обл., Индо-Малайская обл.). Показано видовое богатство дятлов vs общее богатство орнитофауны, доля в нём дятлов в % (3 последовательных столбца после мест наблюдений, дальше — источник и тип леса). Источник. [63], table 1.](http://www.socialcompas.com/wp-content/uploads/2021/11/risu10b.jpg)
б. В тропических лесах (12 мест наблюдений в разных локациях. Строки сверху вниз: Неотропическая обл., Афротропическая обл., Индо-Малайская обл.). Показано видовое богатство дятлов vs общее богатство орнитофауны, доля в нём дятлов в % (3 последовательных столбца после мест наблюдений, дальше — источник и тип леса). Источник. [63], table 1.
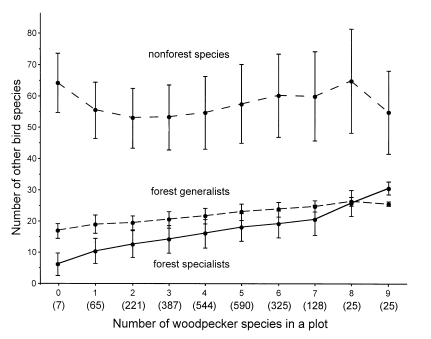
в. Корреляция разнообразия разных составляющих орнитофауны (У) с видовым богатством дятлов, данные по постоянным площадкам, использованным при составлении атласа гнездящихся птиц Польши. Чёрточки — стандартная ошибка средней, цифры в скобках у оси абсцисс — число площадок с данным числом видов дятлов. Снизу вверх: лесные виды — специалисты, лесные виды-генералисты, нелесные виды птиц. Источник. [47], fig.5.
Показана комплементарность участия в этом процессе видов с альтернативными жизненными стратегиями, и разных компонент популяционной системы одного вида (постоянное гнездовое население vs «подвижный резерв популяции») в этом процессе. Чем ниже разнообразие видов дятлов, устойчиво обитающих на данной территории, и ниже численность видов-специалистов полярных стратегий («собирателей» и «долбильщиков»), тем сильней равновесие между сторонами их биоценотической активности сдвинуто вправо, учащая риск появления прорывов полога: «малых» — прореживаний кроны после нападения филлофагов и крупных — окон от усыхания стволов после нападения ксилофагов. Меньшее разнообразие видов дятловых одновременно тормозит метаболизацию мёртвой древесины, образующейся в этих «прорывах» и «окнах», и поднимает риск превращения их в чаги заражения соседних спелых древостоев, приводя к омоложению и гомогенизации выживших. И наоборот: большее разнообразие видов дятловых, с преобладанием специализированных в орнитокомплексе, поддерживает устойчивость более разновозрастных древостоев, с более сложной ярусно-оконной динамикой.
Понятно значение для того и другого именно разнообразия дятлов, а не отдельных их видов: больше их выпало из орнитокомплекса от антропогенного изменения древостоя и прочих причин, тем больше вероятность образования каких-то видов «прорывов» без компенсации. И наоборот, чем большее число видов обитает совместно, одновременно обрабатывает разными способами разные микростации, тем сложнее, мелкомасштабней возникающие прорывы и окна и, главное, они меньше по площади и равномерней в массиве.
СПИСОК ЛИТЕРАТУРЫ
1. Mikusiński G. Woodpeckers: distribution, conservation, and research in a global perspective // Annales Zoologici Fennici. 2006. – V.43. – P. 86-95.
2. Бутьев В.Т., Фридман В.С. Большой пёстрый дятел. Сирийский дятел. Средний пёстрый дятел. Белоспинный дятел. Трёхпалый дятел.// Птицы России и сопредельных регионов. Том Совообразные-Дятлообразные. – М.: Т-во научн.изд.КМК, 2005. – С.328-353, 360-397, 423-434.
3. Фетисов С.А. О гнездовых дуплах и эволюции гнездостроения у дятлов Picidae: Контакты дятлов с другими животными, встречающимися в их дуплах// Русск. орнит. журн. – 2018. – Т.27. – №1570. – С.817-839
4. Wesołowski T. “Lifespan” of woodpecker-made holes in a primeval temperate forest: A thirty year study// Forest ecol. manag. – 2011. – V.262. – №9. – Р.1846-1852.
5. Pakkala T., Tiainen J., Kouki J. The importance of nesting cavity and tree reuse in the three-toed woodpecker Picoides tridactylus in dynamic forest landscapes. // Ann. Zool. Fenn. – 2018. – V. 54. – №1–4. – Р.175-191.
6. Remm J., Lõhmus A. Tree cavities in forests–the broad distribution pattern of a keystone structure for biodiversity// Forest Ecol. Manag. – 2011. – V.262. – №4.
Свежие комментарии