
Моллуккские ибисы в Сиднее
Резюме. Показана контрпродуктивность использование доместикации (дестабилизирующего отбора по Д.К.Беляеву) для понимания урбанизации «диких» видов птиц и млекопитающих, поскольку основано на ложной аналогии. Отчасти сходный результат в том и другом случае достигнут изменениями противоположного характера в жизненной стратегии, поведении, когнитивных особенностях, отношении к человеку и пр. аспектах «естественной истории». «Гены индивидуальности» DRD4 и SERT, при урбанизации находящиеся под позитивным отбором, не участвуют в доместикационных изменениях генома.
Урбанизация движется стабилизирующим отбором, закрепляющим изменения популяционной системы вида, складывающиеся на территории города (в сторону лабилизации отношений) под «давлением» средового стресса, рождённого нестабильностью и изменчивостью «кружева местообитаний», по мере успеха процесса расширяющегося на ещё меньшие, более изменённые и менее стабильные биотопы. Доместикация — движущим отбором, приспосабливающим не популяционную структуру, а непосредственно особей к совершенно иному стрессору (страх «диких» особей перед человеком и созданным им искусственным окружением).
Сходства, вводящие в заблуждение
Когда в руках у тебя молоток, любая проблема кажется гвоздём. Что и случилось с концепцией дестабилизирующего отбора Д.К.Беляева (1983). Она объясняет каскад морфологических, гормональных и поведенческих изменений, идущих у доместицированных видов, создавших их характерные отличия от диких родичей или предков (Трут, 2007, 2008). При этом под отбором оказываются пулы генов, сильно пересекающихся у этих видов и человека1 (Theofanopoulou et al., 2017). Однотипные изменения фиксируются также у кур Gallus gallus (Agnvall et al., 2015).
Это процесс «запускается» и без воздействия человека, одним лишь исчезновением обычных опасностей. Скажем, у домовых мышей Mus musculus, изолированных в амбаре от хищников, за 2002-2016 гг. появились типичные доместикационные изменения — пегости, укорочение лицевой части черепа, уменьшение абсолютных и относительных длин головы и тела (Geiger et al., 2018). Авторы полагают, что, отлавливая и измеряя мышей, непроизвольно осуществили отбор. Из-за этого беспокойства антропофобные мыши рассеивались из амбара, оставались и размножались там лишь самые ручные, а его недоступность для хищников усиливала эффект. Неконтролируемые генетические процессы, подобные этому, обычны в искусственно разводимых популяциях (Артамонова, Махров, 2006)
Правда, не были исключены альтернативные объяснения происходящего: их предшествующая синантропизация. Мыши уже были комменсалами при заселении Европы, в т.ч. менее агрессивными к сородичам, с ослабленной регуляцией «запуска» тревожно-оборонительных реакций, как и доместицированные виды. Происходящее также можно объяснить дрейфом генов, а преобразования черепа — сменой состава корма сравнительно с популяцией-источником (просыпавшееся зерно и фураж, Geiger et al., 2018).
Следовательно, «доместикацией» объясняют многие внешне сходные с ней явления, без специальной проверки, допустимо ли это, гомологичны отмеченные изменения, экологические или микроэволюционные, идущим при одомашнивании или нет? Bозможно ведь чисто поверхностное сходство, которое, при нынешней популярности объяснения доместикацией родит ложную аналогию. Известный этолог Берндт Хейнрих (1994) писал, что вороны Corvus corax воспринимают окружающий мир не как абсолют, но как отклонение от уже принятого, служащего эталоном для сравнения. Люди также «пляшут от печки», действуя под влиянием прайминга и выбирая т. н. социальное сравнение вместо непосредственной оценки ситуации (Аронсон и др., 2008). Существенные различия между явлениями обычно «невидимы» на первом шаге анализа, но уже на втором требуют разделения сходного: от нас, как и антропоидов, это требует больших интеллектуальных усилий, чем обобщение внешне различного (Мешкова, Федорович, 1996; Зорина, Смирнова, 2006).
Задачи статьи:
а) показав невозможность подобного переноса, очертить существенные отличия урбанизации «диких» видов от доместикации домашних, в т.ч. различия стрессоров, к которому приспосабливаются. В первом случае это наиболее общие характеристики урбосреды — экстремальный уровень гетерогенности, нестабильности и изменчивости городского ландшафта cравнительно с прочими «рукотворными2». Он требует от «городских» птиц быстро менять места гнездования, биотопы кормления, кормовые методы т. д. черты биологии вслед за изменением среды обитания, местными и общегородскими, с развитием способности их прогнозировать, жить, постоянно «прыгая с льдины на льдину» в противоположность устойчивому существованию в «сельских» или «лесных» популяций вида (Фридман, Ерёмкин, 2009: 30-35, табл.3; Sprau, Dingemanse, 2017; Isaksson et al., 2018). Во втором — просто смена реакции на людей, технику, уход за животным с тревожно-оборонительной на дружелюбно-интересующую, восприятие их как части окружения и партнёра по взаимодействию, а не опасности;
б) рассмотреть множественные следствия этих различий для биологии «городских» популяций сравнительно с доместицированными, на их основе предложить эволюционный сценарий урбанизации «диких» видов, объясняющий как различия, так и сходства с доместикацией на одном основании.
Урбанизация как приспособление и формообразование
Содержание
«Давление» урболандшафтов, меняющее местообитания в их зонах влияния — главный «движитель» эволюции современных видов, от сосудистых растений с беспозвоночными до птиц и млекопитающих, города — «арена» микроэволюционных процессов, наиболее значимая в нашу эпоху, почему привлекает всё больше внимания (Johnston, Munshi-South, 2017; Beans, 2019). Урбанизация «диких» видов, в нашей работе птиц и млекопитающих, требует наиболее общего объяснения. «Доместикация» часто используется в этом качестве, из-за успешного применения к внешне сходным явлениям (Evans et al., 2010; Møller, 2008, 2010ab, 2014; Alberti et al., 2017) и вхождения в моду истории «ручных русских лисиц» у образованной публики на Западе (Хейр, Вудс, 2014; Дугаткин, Трут, 2019).
Получается плохо, с массой огрехов методологического плана, когда объясняющему соответствуют далеко не все относящиеся к делу данные, но «неудобные» опускаются или отбрасываются. Как это описано в детективе Гарднера:
«Факты, — сухо сказал Дрейк, — вроде обрывков картинки‑головоломки. Мне платят за то, что я их нахожу, тебе — за то, что ты их складываешь вместе. Если они окажутся не от той картинки, ты всегда сможешь засунуть ненужные туда, где их никто не найдет» («Дело о коте привратника»).
Дрейк — сыщик, добывающий информацию, Мейсон — адвокат, строящий из её «кусочков» теории, выдерживающие прокурорскую критику и убедительные для присяжных. Дальше мы «соберём» эволюционный сценарий урбанизации по историям «отношений» разных видов птиц и разных городских ареалов, как собирают паззл, ничего не выбрасывая и не опуская.
Урбанизация «диких» видов всегда адаптивна. Даже на первой стадии, когда «городских» поселений немного и они «держатся» лишь «подтоком» извне (Лыков, 2009; Лыков и др., 2009; Фридман, Ерёмкин, 2009: 9, 72-73, табл.2; Evans et al., 2010, 2011; Фридман, Суслов, 2018), их существование «расширяет» видовую нишу на ещё один, новый, исторически нетипичный ландшафт. Лабилизируются пространственно-этологическая структура поселений и жизненная стратегия особей (она — как бы «отпечаток» отношений, образующих эту структуру — социальных, территориальных и биотопических связей, в перемещениях и взаимодействиях особей), их социальные связи, привязанность к территории и биотопу
а) делаются всё изменчивей, при
б) всё большей зависимости силы связи от «качества» локальной среды, их всё меньшей инерционности (т. е. бывшей прежде зависимости от состава соседей при прошлом размножении, от территории и биотопа тогда);
в) всё более лёгком «переключении» в зависимости от контекста, включая полное прекращение «связи с…» и перемещения в другое место (Фридман, Ерёмкин, 2009; Фридман и др., 2016).
Ещё сиднейские ибисы

Следствия этого:
1) более долгая жизнь «городских» особей, большая «дробность» распределения репродуктивного потенциала между попытками размножения, большая вероятность «отказа» от размножения в каждой из них, при неподходящих условиях (Miranda, 2017; Sepp et al., 2018; Sayol et al., 2020);
2) жизненный цикл городских популяций меняется в сторону «более K», тогда как исходные — «более r-3» (Lowry et al., 2013; Sol et al., 2016, 2018; Sayol et al., 2016), если использовать популярную в эволюционной экологии дихотомию «r-» и «К-видов» (или «-отбора»);
3) Репродуктивный выход «городских» популяций значимо возрастает даже у крупных видов, чувствительных к изменениям местообитаний, как у совиного лягушкорота Podargus striatus в пригородах Мельбурна (Weaving et al., 2016), хохлатой желны Dryocopus pileatus — Сиэтла (Tomasevic, Marzluff, 2018, 2020), перепелятника Купера Accipiter cooperi — Альбукерке (Milsap, 2017) и пр. Так, радиопрослеживание 20 хохлатых жёлн Dryocopus pileatus в 9 районах Сиэтла с разной степенью урбанизации (сохранно от 5% до 95% прежнего лесного покрова) показывает возможность постоянного обитания и успешного гнездования вида при сохранении всего лишь 20% прежнего лесного покрова, но обязательно с крупными мёртвыми деревьями, используемыми для гнездования и ночлега резидентов. Даже в таких условиях самые крупные дятлы США не «жмутся» в оставшихся лесных «островах». Используя их как «базу операций», они чем дальше, тем больше разными способами осваивают селитебную зону: выходят в жилые кварталы, где имеются лишь куртины деревьев, там пользуются кормушками и др. (Tomasevic, Marzluff, 2018, 2020).
«Выигрыш» лысух от освоения Лодзи — в улучшении физического состояния особей: масса птиц (с поправкой на размер) и содержание гемоглобина растут от пригородов к центру. Также снижается гибель гнёзд, вероятность вылупления хотя бы одного птенца у «городских» лысух Fulica atra выше на 29-33%, при 4хкратной разнице репродуктивного выхода с пригородами. В отличие от большинства исследований, не отмечено разницы в выживаемости, связанных с урбанизацией (Minias, 2016).

Рост репродуктивного выхода показан Б.Д.Курановым (1986) для садовой камышёвки Acrocephalus dumetorum в парках г.Томска (доля успешных гнёзд в городе 72.9%, слётков на успешную попытку размножения – 5.09±0.14 vs 63.8% успешных гнёзд, 4.41±0.16 слётка на успешную попытку размножения в пригороде. На 10-20% увеличен репродуктивный выход в популяциях Томска и Северска птиц-дуплогнёздников: горихвостки-лысушки Phoenicurus phoenicurus, большой синицы, скворца Sturnus vulgaris сравнительно с пригородными и «лесными». Это достигнуто за счёт значимо большей успешности выкармливания и насиживания (все виды, кроме пеструшки), даже при меньшей кладке «городских» птиц (большая синица и пеструшка). У «городских» горихвосток вырастает и размер кладки (Куранов, 2009, табл.2).
Рост подвижности 3-х типов связей, формирующих пространственно-этологическую структуру популяции — социальных, биотопических, территориальных — снижает риск исчезновения вида из города вследствие негативных изменений «кружева» местообитаний (Фридман, Ерёмкин, 2009, табл.3). Или из региона — вследствие центробежного распространения таких изменений в нём урбанизацией, и подъёма динамических популяций (Фридман и др., 2016; Фридман, 2018, разд.6).
Следовательно, первый «выигрыш» от урбанизации (шире, от появления динамических популяций, занимающих разные типы «рукотворных ландшафтов») — рост резистентности к средовым изменениям, исходно неблагоприятным для вида, а то и непереносимым (Фридман, Суслов, 2018; Dukatez et al., 2020), в пределе — обращение их во благо: заселение биотопов, первоначально избегаемых, с достижением там большей численности, чем в «видотипичных». Второй — репродуктивный выход городских популяций повышается из-за их превращения в «более К-стратегов»: выживаемость возрастает, среднеожидаемая продолжительность жизни (СОПЖ) удлиняется «в обмен» на снижение величины кладки и выводка4, на подъём доли неразмножающихся птиц («подвижный резерв»), хотя и в меньшей степени. См. Фридман, Ерёмкин, 2009, табл.5; мета-анализы Chamberlain et al., 2009; Vincze et al., 2017; Sepp et al., 2018; Kettel et al., 2018.
Важный фактор подъёма выживаемости в городах особенно крупных и/или охотничьих видов — запрет разряжать оружие в пределах городской черты, не говоря об охоте, чем птицы и пользуются. Индивидуальное мечение 41 канадской казарки Branta canadiensis в Чикаго (2014-2016 гг.) показало, что зимовка здесь даёт больше шансов на выживание, чем перелёт в однотипные ландшафты вокруг. Все 35 «городских» птиц выжили, из ушедших из города — лишь 48%; 70% казарок вернулись в Чикаго до начала осенней охоты и 85% ни разу не покидали его после открытия сезона в середине октября (Dorak et al., 2017). Часто урбанизация видов-жертв начинается раньше, чем их хищников и специализированных конкурентов, город оказывается «островом безопасности», что сильнее подстёгивает освоение. Впрочем, дневные хищные птицы, совы и хищные млекопитающие (лисы, койоты, разные куньи) урбанизируются не хуже прочих (Лыков, 2012; Фридман и др., 2016; Cooper et al., 2020), не говоря о таких разорителях, как белки, сони, крысы, опоссумы. Так, учащается разорение гнёзд голубых сорок Cyanopica cyana в г.Благовещенске белками Sciurus vulgaris, подкармливаемыми посетителями в тех же парках, из-за чего они вынуждены менять места гнездования (Дугинцов, 2018).
Третий фактор — климатический: более мягкие зимы, обусловленные городскими «островом тепла», и растительностью, смягчившей негативное воздействие «волн жары» летом на рост птенцов (в сравнении с «лесными» популяциями, данные по большим синицам Parus major, Pipoly et al., 2020). Зимняя подкормка также вела к более раннему размножению и увеличивала репродуктивный успех на участках, где велась, сравнительно с контрольными, хотя её прекратили за 6 недель до начала репродукции (Robb et al., 2008). Ещё более значимо расширение кормовой базы фито- и энтомофагов из-за увеличения продуктивности при умеренном нарушении природных сообществ (Жигарев, 2004, 2007; Фридман, 2018).

Красношейные поганки Podiceps auritus во время насиживания. Большой пруд
Екатерининского парка г.Пушкина. 25 мая 2018. Фото И.В.Столяровой
В них также повышена доля взрослых и опытных птиц в сравнении с первогодками, облегчая выработку инноваций и распространение их путём социального обучения там, где это возможно5, способствуя более раннему гнездованию как у коротко-, так и долгоживущих видов (Lowry et al., 2013; Sepp et al., 2018). На территории города оно уже обеспечено более ранним ходом весны, часто также более ранним пиком обилия голых гусениц (Chamberlain et al., 2009; Deviche, Davies, 2014; Isaksson et al., 2017), эффект этих двух факторов суммируется.
Так, наблюдения 2011-2015 гг. показывают, что урбанизированные ястреба Купера вытесняют «сельских» в с.-в. Альбукерке (штат Нью-Мексико). Отслеживая вновь вылетевших самок с радиопередатчиками, обнаружили а) региональные популяции сами по себе размножались достаточно успешно, но б) «городские» особи вытесняли их с гнездовых участков, в т.ч. потому что были зимующими, в) экспансия колонистов из города в 30 раз превышала обратный поток. Что говорит об исключительной успешности городской популяции: иначе бы она была не «источником», а «стоком». Успех «городских» особей также поддерживается массовым кормом, отсутствующим вовне — размножившимися с 1980-х гг. белокрылыми горлицами Zenaida asiatica. Их многочисленность позволяет ястребам зимовать, т. е. гнездиться раньше, а, значит, успешней (Milsap, 2017). См. также данные по ястребам, освоившим города разных континентов: чёрно-белому A.melanoleucos в Кейптауне, хохлатому A.trivirgatus в Тайчунге (центральный Тайвань), перепелятнику A.nisus в Люблине, Лодзи, других польских городах (Biaduñ, 2006; Janiszewski et al., 2012; Lin et al., 2015; Suri et al., 2017).

Третий «выигрыш» — устойчивость к островному эффекту, тем более гибельному исходным популяциям, чем консервативней их жизненная стратегия. Все три возрастают тем больше, чем дальше продвинулась урбанизация вида, чем более изменённые (и ближние к центру города) территории он заселит (Фридман, Ерёмкин. 2009; Лыков, 2009; Лыков и др., 2009). Т.е. урбанизация увеличивает приспособленность как «городских» особей, так и «оставшихся» в исходных биотопах.
Действительно, первые поселения вида в городе концентрируют в основном птиц «подвижного резерва», проигрывающих конкуренцию за территорию и партнёра (т. н. floaters, Winker, 1998). Особенно молодых: ювенильная социальность способствует приживанию, а городской «остров» тепла, подходящие биотопы или источники корма обеспечивают зимовку, стимулируя переход ко всё большей оседлости — важный драйвер урбанизации перелётных видов (Баник, 2012; Фридман и др., 2016).
В исходных популяциях они оттеснены в «худшие» местообитания — сильней трансформированные, с большей средовой стохастичностью, следующей отсюда худшей доступностью корма и пр. (Penteriani et al., 2011; Lenda et al., 2012). Но именно поэтому они «выиграют» в городе, где доминируют подобные биотопы, особенно с учётом большего обилия ресурсов и лучшей выживаемости самих птиц, а часто и кладок. Действительно, у больших синиц и лазоревок молодые более инновативны, у них более вероятно социальное обучение, обеспечивающее «культурную передачу» новых навыков. Это согласуется с гипотезой, что птицы «вкладываются» в такие потенциально затратные формы поведения, как инновации с их «культурной передачей», если их текущий статус неудовлетворителен. В поддержку этой идеи, у больших синиц проблемные задачи лучше решают особи, проигрывающие конкуренцию на местах кормления, т. е. низкоранговые члены стай, у лазоревок в экспериментальной ситуации подчинённые более склонны к воспроизведению нового навыка (Aplin et al., 2013, 2015; Lefebre, Aplin, 2017). У грызунов самые инновативные среднеранговые зверьки: большинство сообщества, уходящее за счёт изменений поведения от агрессии доминантов и получающие сравнимую с ними свободу передвижения, исследования, кормления и пр. (Мешкова, Федорович, 1996: 78-86)

Действительно, первые поселения в урболандшафте созданы преимущественно «худшими» особями региональных популяций (Фридман, Ерёмкин, 2009: 47-53). Это легко объясняет общий «сдвиг» жизненной стратегии в городских популяциях — больший вклад в самосохранение для будущего размножения в лучших условиях, активный их поиск, уменьшенный вклад в репродукцию «здесь и сейчас» (Sol et al., 2018; Sepp et al., 2018). В «природных» популяциях он отличает особей, проигрывающих конкуренцию: уязвимые к социальному стрессу, они более устойчивы к средовому, отчасти даже выигрывают от него (Фридман, Ерёмкин, 2009, разд.II.4-6).
Судьба птиц из «резерва» там незавидна: больший риск гибели, худшее физическое состояние, больший стресс и минимальные шансы на размножение. Наибольший успех связан с занятием «вакансий» владельцев территорий и/или их брачных партнёров, «освобождающихся» с их смертью или переселением, но и здесь преимущество за резидентами тех же группировок (Penteriani et al., 2011; Lenda et al., 2012). В основном они гибнут, не реализовав репродуктивный потенциал, за единственным исключением попадания во временные высокопродуктивные биотопы, ещё не заселённые данным видом, т. к. возникли недавно. Их подток уменьшает риск вымирания изолированных поселений от случайных причин (Robbles, Cludad, 2017, 2018).
Размножение там идёт интенсивно, однако находка подобных участков в природных ландшафтах — событие редкое, непредсказуемое для птиц из «резерва»; случайный рыск здесь неприемлем, т. к. крайне опасен, почему переселения внутри популяций направленные (Фридман, 2016). Урбанизация решает эту проблему. «Метаболизм» и рост городов «производят» такие участки во всё большем числе, на большей площади etc. Одновременно возникает градиент изменённости местобитаний, целенаправленно «наводящий» на них птиц из «подвижного резерва» (Фридман и др., 2016; Фридман, 2018).
«Движение» потенциальных урбанистов по городскому градиенту «вверх», в сторону больших уровней средового стресса продолжается и на территории собственно города, пока урбанизация вида не завершится. Благодаря этим перемещениям все виды при урбанизации «экспонируют» значительную часть особей в новых условиях, не только генералисты, как часто считается (Sol et al., 2016; Sayol et al., 2016). Их поселение в урболандшафте «убивает двух зайцев»: повышает приспособленность как ранее «худших» особей, так и среднюю по популяции.
Что повторяется раз за разом в заселяемых птицами городах, например, у сороки Pica pica в разных городах Палеарктики, от Дании и Франции до Приамурья (Лыков, 2017, табл.1); у вяхиря Columba palumbus в Калининграде; Бресте и других городах Белоруссии6 (Сахвон, 2016; Sakhvon, Köver, 2020); Львове и других городах Галиции (Фридман, Ерёмкин, 2009); в городах Финляндии (Fey et al., 2015); чёрного дрозда Turdus merula в разных городах Европы; лысухи в Лодзи, Калининграде, Берлине; камышницы Gallinula chlorophus в Москве, Берлине, Калининграде (Otto, Witt, 2002; Березовиков, Таболина, 2014; Лыков, 2015) и т.д. «Истории урбанизации» однотипны у видов, очень разных экологически и друг другу неродственных (Фридман, Ерёмкин, 2009, табл.2; Evans et al., 2010; Lowry et al., 2013; Фридман и др., 2016).
От урбанизации «выиграет» и вид как целое. «Нашлась» новая ниша для особей, раньше лишь увеличивавших груз элиминации (=снижавших приспособляемость популяции согласно дилемме Холдейна, Suslov, 2013), расширяется спектр местообитаний, т. е. больше возможности «попадания» родившихся там особей разного «качества» (с разной индивидуальностью, жизненными стратегиями и пр.) в участки с условиями существования, наиболее подходящими именно им. Что достигается направленными переселениями птиц из «подвижного резерва» популяции между поселениями, существующими на разных участках этого спектра (Фридман, Ерёмкин, 2009; Фридман и др., 2016).

Источник: Резанов А.Г., 2016. Зимовка лутка Mergellus albellus на реке Москве в Коломенском: распределение и кормовое поведение.


Зимовка северных нырковых уток на тёплых Курьяновских сливах в Москве
Это т. н. «сортировка» особей разного «качества» (с потенциями формирования разного поведения и экологических предпочтений, задаваемых чаще всего одной из двух или нескольких альтернатив, существующих в данной популяции) между более или менее подходящими им участками местообитаний, когда те формируют пространственный градиент (-ы) средовых изменений. Переселенцы — в основном птицы «резерва» (с преобладанием молодых); с небольшой, но значимой долей взрослых владельцев территорий, успешно размножавшихся там. См.данные по становлению урбопопуляций тетеревятника в Гамбурге (Rutz, 2008); золотоголового дроздового певуна Seiurus aurocapilla в пригородах Weston, штат Массачусетс (Morimoto et al., 2012); черноголовой гаички P.atricapilla в Campose, пров.Альберта, Канада (Perkins, 2017), и обзоры Kristan et al., 2007; Penteriani et al., 2011; Lenda et al., 2012.
Из градиентов в большинстве регионов наиболее выражен городской, вдоль него особи «сортируются» между возникающей урбо- и региональной популяцией. Урбофобы и их противоположность — потенциальные урбанисты могут появиться в любой из двух. Благодаря «сортировке» те и другие с высокой вероятностью попадут в наиболее подходящую для себя среду обитания (Фридман, 2012, 2018; Фридман и др., 2016). Направляясь сигнальным полем отдельных группировок, «сортировка»
1) делает расселительные процессы в популяции направленным перемещением особей вместо случайных блужданий;
2) Благодаря ей популяции вида на разных «сегментах» ландшафтных и географических градиентов (a) спаяны в целое — вид как систему популяций с ареалом, обнимающим широкий спектр местообитаний на большой территории (т. н. «вид Вавилова»), (b) по той же причине растёт разнородность между отдельными популяциями, они чем дальше, тем больше дифференцируются друг от друга, приспосабливаясь к различающимся местным условиям — но не каждая по отдельности, а за счёт общего процесса сортировки. Предположенная сперва на основании косвенных данных (Фридман, Ерёмкин, 2009, разд.II.3; Фридман, 2012), сегодня она показана строго (многоголосый пересмешник Mimus polyglottus, Stracey, Robinson, 2012; кроличий сыч Athene cunicularia, Rebolo-Ifrán et al., 2015; перепелятник Купера, Milsap, 2017; большая синица, Charmantier et al., 2017; Caisergues et al., 2018; озёрная чайка Larus ridibundus, Indikiewicz et al., 2018).
Точно также дивергенция приспособлений и «естественной истории» в целом между альтернативными популяционными системами вида в регионе, «городскими» и «сельскими» или, шире, динамичными и консервативными, имеет тенденцию увеличиваться по ходу его урбанизации, когда «городской градиент» делается длинней и/или круче. «Сортировка» одновременно и связывает между собой разнотипные популяции, и — благодаря неслучайным переселениям особей в обе стороны «вдоль» градиента средовых изменений — усиливает экологические, поведенческие, генетические отличия между ними (Фридман, 2012; Evans et al., 2012; Møller et al., 2012; Фридман и др., 2016; Minias et al., 2017; Isaksson, 2018).
«Сортировка» переселенцев между городскими vs исходными биотопами повышает устойчивость «городской» популяции в самый важный период — начало урбанизации, когда птицы успешно живут и размножаются в городе, но большая часть молодых уходит вовне, где запечатлевают будущий гнездовой район, почему вряд ли вернутся в урболандшафт (Фридман, Ерёмкин, 2009, разд.II.3; Фридман и др., 2016). Тогда «городская» популяция поддерживается лишь притоком птиц, вытесненных из исходной, в чём нет ничего необычного: во многих местообитаниях из ряда занятых видом в данной местности поддерживаются притоком птиц из «резерва», а не размножением резидентов. Для сохранения редких видов в такой ситуации успешное «осаждение» их в нужных местах проигрыванием вокализаций, подстановкой макетов и пр. более значимо, чем охрана самих гнездовых группировок (Robles, Cuidad, 2017, 2020; Фридман, Суслов, 2018).

Двусторонняя «сортировка» особей между городской и региональной популяцией увеличивает первую за счёт репродукции второй; обе пополняются особями с особенностями индивидуальности и поведения, заставляющими избегать биотопов места «исхода», но благоприятными в местах закрепления. Фактически она «пролагает дорогу» отбору, поддерживая высокую скорость приспособительных изменений при минимуме элиминации: и выживаемость птиц в городских популяциях, и их численность возрастают сравнительно с исходными.
Важно отметить, что первым по ходу урбанизации фиксируется подъём пластичности популяционных и социальных реакций, связанных с прохождением жизненного цикла и окружением особи в этом процессе: выбором поселения, территории и соседей внутри него, местообитаний для гнездования и кормления внутри территорий, факторов, запускающих смену поведенческих стереотипов в прежнем месте обитания и/или переселение в новые.
Что вызывается растущим «давлением» городских ареалов на местообитания вида в регионе, и проявляется в их фрагментации на всё меньшие и всё более изолированные «острова», трансформации их «с краёв», т. е. изменением «кружева» местообитаний, с одной стороны, всё более непредсказуемым для птиц, с другой — в неблагоприятную сторону. Вовне это проявляется в лабилизации жизненной стратегии вида, исходно существенно более консервативной, если оценивать местом на граденте «лабильность-консерватизм», впервые фиксирующемся ещё до освоения собственно урбосреды, в пограничных местообитаниях вида в регионе, начавших изменяться «только что» под влиянием городских ареалов (сопряжённо растущих в составе агломерации), «дошедшим» до них буквально недавно (Фридман, Ерёмкин, 2009; Фридман и др., 2016; Фридман, Суслов, 2018)..
«Фронтир» расползающегося «давления» городских центров — стартовая позиция урбанизации вида (отнюдь не биотопы, занятые уже внутри города по ходу последней). Рост пластичности индивидуальных характеристик, на который натуралисты традиционно обращают больше внимания, происходит сильно потом — когда популяции с изменённой пространственно-этологической структурой успешно освоят урболандшафт, проникнут вв нехарактерные местообитания, в которых удерживаются лишь благодаря лабилизации. И через некоторый период жизни в них появляются те поведенческие инновации и экологические изменения, которые полагают признаком успешной урбанизации, тогда как они её следствие, и достаточно отдалённое (Фридман, Ерёмкин, 2009; Фридман и др., 2016; Фридман, Суслов, 2018).

Отбор, в свою очередь, делается стабилизирующим: он «подгоняет» фенотип к новой норме, заданной вышеописанной сменой пространственно-этологической структуры популяций и вызванной ею коррекцией моделей поведения на уровне особей. Если трансформация обоих исключительно быстра (годы), то «подгонка» отбором существенно медленней (первые десятки лет). Изменения морфофизиологии и генных вариантов при урбанизации потому и запаздывают (часто значительно), что стабилизирующий отбор в городских популяциях как бы проводит тюнинг для птиц, успешно освоивших урбосреду и в ней благополучных. Для этого хватит лабилизации популяционной системы (вместе с имеющейся пластичностью организмов, Miranda, 2017; Liker, 2020), а дальше происходит «доводка» отбором. См. ниже и Суслов, 2014; Фридман и др., 2016; Фридман, Суслов, 2018.
Иными словами, города «концентрируют» особей с определённым поведением (его отличия от исходной популяции бывают разного «масштаба»: от жизненной стратегии в целом до отдельных стереотипов и особенностей индивидуальности), отличающейся физиологией, генотипами и пр. со всего региона. И именно тех, что в урбосреде получают преимущество над «дикими типом» (Müller et al., 2013ab; van Dongen et al., 2015), тогда как вовне их приспособленность снижена, см.данные по чёрному дрозду (Evans et al., 2009) и кроличьему сычу (Rebolo-Ifrán et al., 2015; Luna et al., 2019). И наоборот, в регион перемещаются птицы, чьё поведение, экологические предпочтения, генотип имеют преимущество в «сельских» или «лесных» популяциях вида. Направленная сортировка особей по поведенческим потенциям и/или основанным на них альтернативным стратегиям идёт и внутри «городской» популяции, между разными мозаиками урболандшафта и, соответственно, районами города (сизарь C.livia, большая синица, Фридман, Ерёмкин, 2009, разд.II.3-4; большая синица, Sprau, Dingemanse, 2017 и др.).
На первых порах освоения урболандшафта обмен особями с регионом не слабеет, но даже усиливается, а дифференциация «городских» особей происходит за счёт роста связности «материнской» и «дочерней» популяций. Когда вторая достаточно выросла, а урбанизация заметно «продвинулась» вперёд (в т.ч. птицы заняли настолько изменённые местообитания, что вне города просто отсутствуют), она отделяется, всё больше пополняясь собственным размножением, обретает независимую историю и судьбу (Фридман, Ерёмкин, 2009; Фридман и др., 2016; Miranda, 2017; Фридман, 2018; Isaksson, 2018). Сейчас этот вывод подкреплён данными об их генетической дивергенции от исходных (серый юнко Junco hyemalis Rasner et al., 2004; обыкновенная пустельга Falco tinnunculus, Rutkowski et al., 2006; чёрный дрозд Evans et al., 2009; большая синица Björklund et al., 2010; лысуха, Minias et al., 2017).

Египетские цапли кормятся на рыбном рынке в Керале. Источник Rosnath, Palatty, 2017
Больше того, в городских популяциях местных видов (чёрный дрозд, большая синица, чёрный лебедь Cygnus atratus, кроличий сыч) и закрепившихся экзотов (наполеонов бархатный ткач Euplectes afer на Пиренеях; лесная завирушка Prunella modularis7 в Новой Зеландии сравнительно с английскими птицами) избирательно накапливаются генные варианты, облегчающие приспособление к наиболее общим характеристикам урбосреды — максимальным гетерогенности ландшафтной структуры, нестабильности каждого участка мозаики «здесь и сейчас» и скорости его изменений на более длинных временных интервалах. Среди генов, меняемых урбанизацией «диких» видов птиц и млекопитающих8, наиболее важны 3: ген серотонинового транспортера SERT, дофаминового рецептора D4 DRD4 и связанный с биоритмами ADCYAP1 (adenylate cyclase activating polypeptide 1 — кодирует белок, регулирующий синтез мелатонина – гормона регулятора суточных и сезонных ритмов активности, усвоения пищи, обмена веществ и др.). Они показывают максимум направленных изменений (накопление «городских» вариантов SNP в позициях, соответствующих функциональной части гена, экзонах и микросателлитах), почему считаются кандидатными генами урбанизации (Müller et al., 2013ab, 2014, 2017).
Первый влияет на стрессоустойчивость индивида, т. е. способность действовать в условиях опасности и иных травмирующих воздействий, вроде фрагментации местообитаний, их рассечения коммуникациями и пр9., а также на реактивность, т. е. выбор и амплитуду ответа. Так, максимальное выражение антихищнического поведения у больших синиц — птица шипит и вертит головой, отчасти создавая впечатление змеи, особенно при нахождении в дупле. Timm et al. (2019) показали, что наличие или отсутствие этой реакции на опасность у данного индивида коррелирует с изменениями в экзоне 1 гена SERT (SNP 187), объясняющими 16% дисперсии поведенческого ответа, но не связано с какими-то полиморфизмами гена DRD4.

Второй влияет на когнитивные функции особи, связанные с реагированием на новое и неизвестное в её окружении (объекты, события, процессы), требующее исследования, классификации, и выбора способа реагирования, хотя и потенциально опасное, исходно пугающее. Именно при освоении городов (в меньшей степени — новых для вида территорий при успешных инвазиях, ещё меньше в других «рукотворных ландшафтах») частота встреч с ними резко растёт сравнительно с исходными биотопами, достигая абсолютного максимума. Носители «городских» вариантов DRD4 отличаются лучшей способностью к категоризации и большим объёмом оперативной памяти, как и способности их задействовать в условиях стресса, более оптимальным распределением времени между задачами. В природных популяциях (сейшельская камышовка Acrocephalus sechellensis) и инвазивных популяциях (волнистый астрильд Estrilda astrild) связь черт индивидуальности с вариантами гена DRD4 и SERT может отсутствовать: урбанизация и/или инвазия на новые территории её усиливает, возможно, и создаёт de novo (Johnston, Munshi-South, 2018; Liker, 2020; Silva et al., 2020).
Среди них варианты гена постсинаптического дофаминового рецептора D4 (DRD4), консервативного у птиц и млекопитающих, включая человека: связан с приоретизацией внимания и объемом рабочей памяти, что (среди прочего) определяет лучшую переключаемость и многозадачность. В городских популяциях больших синиц избирательно накапливаются варианты DRD4, обеспечивающие лучшую категоризацию и дифференциацию ситуаций, оптимальное распределение времени между конкурирующими задачами, а самих индивидов — между участками урболандшафта с разным характером беспокойства (Fidler et al., 2007; Müller et al., 2013b; Riyahi et al., 2015, 2017; Sprau, Dingemanse, 2017). Сходные изменения связаны с урбанизацией или инвазией на новые территории у других изученных видов (Müller et al., 2013ab, 2014, 2017, 2019; van Dongen et al., 2015; Holtmann et al., 2016).

Молодая прудовая цапля кормится выброшенной рыбой под цапельником в Керале. Источник Rosnath, Palatty, 2017
В городских популяциях тех же видов избирательно накапливаются и варианты гена серотонинового транспортера SERT. Это определяет большую устойчивость к любым стрессорам: среди прочего уменьшается дистанция взлёта. SERT реагирует на урбанизацию быстрей всего (Müller et al., 2013ab, 2014; van Dongen et al., 2015; Holtmann et al., 2016). У дарвиновых вьюрков Geospiza fortis и G.fuliginosa на о.Санта-Крус урбопопуляции отличаются эпигенетически, степенью метилирования разных участков ДНК: морфологические отличия от исходных популяций есть лишь у первых. Не найдено различий в изменчивости числа копий генов (англ. Copy number variation) между популяциями (McNew et al., 2017).
У «городских» больших синиц метилирование CpG-островов в промоторах генов DRD4 и SERT, также как в «острове», перекрывающем экзон 3 гена DRD4, было повышено на 1-4% во всех тканях. Повышен и общий уровень метилирования во всех локусах. Исследовались птицы из городских парков Барселоны vs двух районов природных ландшафтов вокруг. Промотор гена SERT представляет собой бедную по CpG область; метилирование одного динуклеотида CpG, расположенного в 288 п.н. от сайта начала транскрипции, коррелировало уровнем исследовательской активности «городских» птиц. Также генотипы полиморфизма SNP234, локализованные в промоторе SERT, значимо коррелируют со стремлением к новизне, её поиском и исследованием (противоположность неофобии) в неволе, и аллель, усиливающий эту активность, чаще встречается у «городских» птиц. Авторы полагают, что генетическая изменчивость и вариации в метилировании гена SERT существенны как подоснова индивидуальности у больших синиц (Riyahi et al., 2015).
В это же время значительные различия в индивидуальности между большими синицами Барселоны и природных территорий вокруг (выше смелость, интенсивней исследование новых объектов) оказались не связаны с частотами генотипов полиморфизма SNP830 в гене DRD4, хотя эти последние также значимо различались у «городских» и «лесных» синиц. Также отмечено разрушение поведенческого синдрома — исследовательская активность коррелирует со смелостью у «лесных», но не «городских» птиц (Riyahi et al., 2017), как это вообще происходит при урбанизации вида (см.далее). Однако данные по искусственному отбору на соответствующие типы поведения (Fidler et al., 2007), также как по другим городам Европы (Müller et al., 2013b) показывают значимую связь индивидуальностей особей с этим и другими полиморфизмами в DRD4.
Депрессия матери в пренатальном периоде меняет паттерн метилирования промотора SERT у плода. Его уровень метилирования также связан с плохим обращением в детстве. Уровень метилирования DRD4 и SERT также отрицательно коррелирует с развитием симптомов синдрома дефицита внимания и гиперактивности. На протяжении жизни у однояйцевых близнецов накапливаются различия в метилировании генов SERT и DRD4, отражая накапливающееся несходство их биографий и жизненных обстоятельств (Riyahi et al., 2015).
Как и в случае с человеческим детством, вырастание в экстремальной среде создаёт «отпечаток» в виде метилирования генов, меняющего уровень их экспрессии в сторону, обеспечивающую первый этап адаптации — интенсификацию функции, «попавшей» под действие стрессоров, чтобы и в этих условиях «жить, ничего не меняя». Фактически это аналог т. н. длительных модификаций, наиболее известных у растений и беспозвоночных. Согласно модели парфорсной эволюции, позже, уже на втором этапе приспособления, уже замещаются генетическими изменениями с противоположным эффектом: обуславливая сдвиг морфологии, физиологии, поведении вида, они создают ситуацию, когда ранее экстремальная среда делается благоприятной (Суслов, 2011; Suslov, 2013). Действительно, метилирование «генов индивидуальности» DRD4 и SERT в «городских» популяциях птиц значимо отличается от такового в исходных и как минимум у больших синиц эти различия связаны с изменением индивидуальности при урбанизации. У них же различие транскриптомов между популяциями связывают с эпигенетическими изменениями уровней экспрессии генов (Riyahi et al., 2015; Verhulst et al. 2016; Watson et al., 2017).
Урбанизация иначе меняет морфологию
Как известно, действующая причина доместикационных изменений — задержка миграции клеток нервного гребня, запускающая каскад изменений на уровне морфофизиологии и поведения; изменения тимуса (ювенализация) объясняют лишь частные аспекты происходящего (Попова, 2015; Уилкинс, 2017). Поэтому они проявляются в пегостях, вислоухости, укорочении морды и т. д. преобразованиях морфологии.
Ничего этого не отмечено при урбанизации. Часто утверждается рост частичного альбинизма и т. д. пегостей у синантропных видов птиц (домовый воробей Passer domesticus, серая ворона Corvus corone cornix, галка Corvus monedula и пр., Ильенко, 1976, цит.по: Корбут, 2018). Однако это модификационные изменения, как хромизм и т. д. аберрации, их частота повышена в Москве сравнительно с малыми городами и посёлками, не «сельскими» популяциями данных видов. Рост частоты аберраций отмечен в период линьки или у молодых, когда часты затруднения с качественным питанием, и снова снижается в дальнейшем. У серой вороны и домового воробья она была снижена в годы подъёма численности, и вырастает при спаде, когда увеличилась доля слётков в составе популяции. Полноценное питание птиц с пегостями при клеточном содержании нормализует окраску после первой же линьки (Корбут, 2018). Немногие дожившие до взрослости аберранты отличаются исключительной осторожностью, по-видимому, в порядке компенсации, т. к. избирательно отлавливают ястреба (Тищенков, 1999; Караваев, 2013). То же отмечено относительно пегостей «городских» чёрных и певчих дроздов в Великобритании (Sage, 1963).
Даже на максимуме концентрации распространение пегостей в популяциях синантропных видов птиц несравнимо с происходящим при доместикации: там окрасочные изменения сопряжены с морфологическими, особенно в форме черепа, и охватывают всю популяцию, делаются обычными. В городских популяциях они — редкие исключения (Лукашик, 2018). Учащения пегостей вовсе не происходит видов-синурбанистов, обитающих на «островах» природных территорий, а не в застройке. У населяющих города вяхирей, тетеревятников, воронов, жёлн обыкновенной и хохлатой, зелёных, малых, белоспинных, больших пёстрых дятлов больше пегостей не становится. Наоборот, в ряде случаев урбанизация сокращает пегости: у юнко, освоивших г.Сан-Диего, голова сделалась темнее, а белые каймы хвоста исчезли10.
Кроме подъёма агрессивности, у «городских» певчих воробьёв Melospiza melodia более развиты бурые пестрины на груди: их больше и меланиновая окраска интенсивней. Одновременно «перестроены» её связи с территориальностью: большая выраженность последней в исходных популяциях связана с меньшим пространством, занятым испещрением, но в «городских» свойственна птицам с самыми тёмными пестринами. Гормональная подоснова испещрения в обоих случаях одинакова: большая развитость вызвана повышенной секрецией тестостерона под управлением гонадотропин-релизинг гормонов, у более испещрённых самцов сильней подъём кортикостерона в ответ на стрессор. Напротив, связи окраски с поведением меняются в разных ландшафтах (Beck et al., 2018).

Молодой барсук устроил нору под могильным памятником на кладбище в Бристоле
Одно из таких изменений прямо противоположно пегостям — это меланизм сизарей и других «городских» видов. Меланисты лучше действуют в условиях неопределённости: они психологически устойчивее, спокойней, менее импульсивны, лучше «дозируют» агрессию и исследовательское поведение по цели. Черночеканная морфа сизаря более адаптивная в условиях высокой плотности колоний, при стратегии «собирателей» и «помоечников», кормящихся поблизости от них во дворах и «пешком». «Дикий тип» адаптивен при разреженном гнездовании и/или при стратегии «летунов» (Ксенц, 1983, 1987, 1990, цит.по: Фридман, Ерёмкин, 2009).

Лань пересекает улицу в Лондоне
Кандидатные гены урбанизации не задействованы в доместикации
Главные доводы в пользу сущностных различий между урбанизацией и доместикацией даёт анализ генов, оказывающихся «под отбором» в обоих процессах: 1) это разные гены; 2) их изменённые варианты, «поддерживаемые» селективным давлением в том или ином случае, вовлечены в разрешение качественно различных проблем приспособления. Следовательно, оба процесса «запускаются» разными стрессорами и «на выходе» создают разные адаптации, что анализируется далее.
Два кандидатных гена урбанизации (DRD4, SERT) важны для оптимизации выбора поведения в проблемной ситуации (задача, критически важная для «городских» птиц, тогда как «сельским» или «лесным» достаточно следовать видовым стереотипам), задействуются в двух разных её аспектах, дополняющих друг друга. «Городские» особи постоянно оказываются в обоих, а отреагирование ситуаций, разрешающее проблему или использующее её на благо индивида, осуществляется при аномально высокой нестабильности, изменчивости, и гетерогенности среды. В этом суть средового стресса в урболандшафтах, их коренное отличие от других «рукотворных», а тем более от природных (Фридман и др., 2016, рис.5).
В урбопопуляциях разных видов показан отбор вариантов обоих генов, направленных на облегчение этой задачи в обоих аспектах (Müller et al., 2013ab, 2014, 2017; van Dongen et al., 2015; Riyahi et al., 2015; Holtmann et al., 2016). Данные изменения идут независимо в разных «городских» популяциях одного вида, образуя гомологические ряды как между ними, так и между разными видами. Направленное накопление «городских» вариантов SERT и DRD4 фиксируется не во всех обследованных урбопопуляциях: в части из них оно запаздывает, подтверждая вывод о том, что генетические изменения там значительно отстают от морфологических, а тем более эколого-поведенческих изменений, поскольку все три связаны в первую очередь с адаптацией общего значения, к урбосреде как таковой, и только попутно — к отдельным существенным для вида воздействиям в ней.

Ген SERT связывают c подавлением стресса, обычно возникающего при многозадачности, необходимости переключения между действиями, столкновении с новым и необычным и пр. SERT — ген серотониновой стресс-лимитации, он помогает легче переносить любой стресс, т.н. серотониновая эйфория, в т.ч. исследовать новое и необычное «вопреки» усилению разнотипного беспокойства и «возмущений» среды. Последнее облегчает существование в ней на каждом этапе её освоения «дикими» видами: начальных, когда это мощный стрессор, сдерживающий расширение ареала «городской» популяции везде, кроме ряда «стартовых» биотопов, и конечных, когда это последнее активно идёт, а особи успешно решают проблемы «городской жизни», но и всё больше эффективно используют её обстоятельства (Riyahi et al., 2015, 2017). Это верно и для «городских» изменений гена DRD4, почему оба они суть адаптации общего значения.
На селекцию SERT урбанизацией отвечает быстрее всего. Дивергенция «городских» популяций чёрного дрозда от «лесных» затрагивает экзонный микросателлит, показывающий признаки положительного отбора в 10 популяциях и 12 исследованных. В городских популяциях T.merula частота основного аллеля сравнительно с реликтовой снижена, а доля гетерозигот повышена, причём специфического «аллеля региона» не выявлено даже среди редких, т. е. это параллельные изменения под действием отбора (Müller et al., 2013a; Garroway, Sheldon, 2014).
Видимо, «городские» аллели обеспечивают большую резистентность к тем (преобладающим в потоке событий вокруг городской птицы) ситуациям, и объектам на которые не надо реагировать. Эти он позволяет избегать ущерба и устойчиво существовать особи «внутри» давки и суматохи городской жизни. Благодаря им «городские» птицы делаются «равнодушны» к вышеописанным составляющим «городского метаболизма», «сельские» при тех же воздействиях чаще всего отлетают прочь (и сильно стрессируются, когда остаются на месте). Значительно меньше он задействован в ситуациях, действительно связанных с «обращением Савла в Павла» — доброжелательностью к объектам, ситуациям и воздействиям, раньше вызывавшим лишь тревожно-оборонительную реакцию.
Поэтому он (вместе с DRD4) вносит вклад в большую смелость как «городских» особей сравнительно с «сельскими» и «лесными» (данные по чёрному дрозду, Müller et al., 2019, большой синице, Timm et al., 2019), так и освоивших новые территории за пределами основного ареала. Скажем, в новозеландской популяции лесной завирушки избирательно накапливаются варианты SERT, связанные с большей смелостью, сравнительно с исходной британской (Holtmann et al., 2016). У людей ген связан с тревожностью, избеганием вреда и другими подобными формами поведения.

Неслучайно DRD4 и SERT — т.н. «гены индивидуальности», дающие «биологическую» подоснову дифференциации альтернативных стратегий внутри популяции. Их разделение, в свою очередь, «оформляет» индивидуальность особей, специализирующихся как «носители» той или иной альтернативы. Именно в городских популяциях их разделение проявляется ярче всего, как и дихотомия «представляющих» их индивидуальностей, вроде «быстрых» vs «медленных исследователей» у синиц, «черночеканных» особей vs сизарей «дикого типа» и пр. (Fidler et al., 2007; Фридман, Ерёмкин, 2009).
Так, в «городских» популяциях больших синиц и черноголовых гаичек значимо преобладают «быстро обследующие» («fast» explorers). Носители этого типа индивидуальности, будучи помещёнными в новое пространство, осматривают его интенсивней (быстрей движутся, чаще перемещаются), но «поверхностней» (чаще возвращаются к уже осмотренному). Ему противопоставляются обследующие медленней, но много тщательней («slow» explorers), преобладающие в исходных популяциях: первые смелей, агрессивней, но и стереотипней, склонны к копированию кормовых методов других птиц, вторые осторожны и робки, но инновативны; кормятся самостоятельно. Проблемные ситуации первые разрешают активным действием, вторые — пассивным переносом (проактивная vs реактивная тактика). См. Sih et al., 2004; Thompson et al., 2018.
Большая интенсивность перемещений, в т.ч. энергозатратных, вроде полётов и перепархиваний, увеличение порывистости каждого из них и зигзагообразности общей траектории прямо связано с лучшей ресурсообеспеченностью в городах (как минимум взрослых особей, у птенцов бывают проблемы с белковым кормом именно в нужный период из-за частых случаев меньшего обилия голых гусениц и другого массового корма в городских биотопах, Seress et al., 2018), резко расширившей возможность «активного бездельничанья». Эти черты индивидуальности наследственны — как и другие анализируемые в данной статье: отбор по ним в 4-х поколениях больших синиц был успешен и усилил их альтернативность (Fidler et al., 2007; Riyahi et al., 2015, 2017; Baugh et al., 2017).
Скорей всего, в урбосреде важны оба варианта, но более выраженные, чем в исходных биотопах (отсюда отбор, производимый урбанизацией). У больших синиц и черноголовых гаичек постепенно вырисовывается связь альтернативы «быстрого» vs «медленного» исследователей с альтернативой между скоростью и точностью решения проблемных задач и задач на социальное обучение (Lefebre, Aplin, 2017). У первых преобладает скорость, у вторых точность, почему внутри города они «сортируются» между разными элементами урболандшафта (Sprau, Dingemanse et al., 2017).

Наследуемость обоих вариантов индивидуальности также возрастает при урбанизации, в т.ч. потому что среди генов, влияющих на индивидуальность животного, наиболее важны кандидатные гены урбанизации SERT и DRD4, почему их также называют «генами индивидуальности». Последнее подтвердилось экспериментально. Использование антагонистов рецепторов дофамина для D1 и D2 метаболических путей (последний включает рецептор D4) показало немедленное изменение индивидуальности астрильдов Estrilda astrild после их кратковременного воздействия. Блокирование D2 уменьшает их социабельность (время, проводимое у зеркала в соответствующем тесте, и двигательную активность перед ним). Экспериментальное увеличение D2 действовало противоположно: повышало подвижность в социальном контексте (у зеркала), но снижало в прочих; D1 — увеличивало время, проводимое у кормушки, и интенсивность питания. Среди прочего эти манипуляции разрушили корреляции, исходно присутствующие у астрильдов: более подвижные птицы менее социабельны (Silva et al., 2020). Именно так разрушает урбанизация т. н. поведенческие синдромы, присутствующие у птиц в исходных популяциях: исчезновение связей между составляющими их качествами позволяет каждой из них при освоении данным видом данного города меняться независимо от всех прочих.
«Городские» варианты обоих генов найдены у разных видов птиц и/или накапливаются в популяциях в популяциях разных городов, отличающихся длительностью существования. Третий ген, ADCYAP1, связан с биоритмами и изучен существенно менее. Скорей всего, отбор его «городских» вариантов вовлечён в снижение перелётности птиц «городских» популяций, особенно урбанизирующихся «через зимовку», и/или в последовательное расширение временных границ репродуктивного периода по мере освоения урбосреды. Возможно, он не включается в урбанизацию оседлых видов и/или видов птиц вне умеренного пояса.
Оба гена у птиц и млекопитающих консервативны, влияние их изменений на поведение неплохо изучено (в основном, правда, на людях врачами), позволяя строить предположения о функции вариантов, накапливаемых в «городских» популяциях отбором. DRD4 — ген постсинаптического дофаминового рецептора D4 задействован в лучшей дифференциации ситуаций и оптимальном распределении времени между конкурирующими задачами.
Однонуклеотидные полиморфизмы (SNP) в третьем экзоне гена DRD4 у людей и других млекопитающих связаны с межиндивидуальной изменчивостью по уровню неофилии. консервативен у птиц и млекопитающих, включая человека: связан с приоретизацией внимания, категоризацией происходящего, объёмом оперативной памяти, что (среди прочего) определяет участие его «городских» вариантов в усилении переключаемости и многозадачности у «городских» птиц (как и «длинных» аллелей гена сравнительно с «короткими» у человека»). Это же верно и для млекопитающих. Так, «городские» койоты Canis latrans в Чикаго отличаются увеличением смелости и агрессии (в т.ч. по отношению к человеку) от «сельских», при снижении индивидуальной повторяемости и росте контекстуальности обоих характеристик, особенно второй. В отличие от «сельских» койотов, изменения смелости и агрессии у одних и тех же «городских» особей не коррелируют между собой (Wush, 2018).
DRD4 жёстче связан с чертами индивидуальности особей у воробьиных Passeriformes, нежели у прочих птиц (однако у ряда видов такой связи не найдено, либо, как у большой синицы, найдена лишь в ряде популяций). В этом качестве он влияет на страх и его противоположность — стремление к новизне и исследовательскому поведению. У воробьинообразных он и эволюционирует быстрей, чем у прочих птиц. Исследованиями индивидуально меченых особей трёх видов — мухоловки-белошейки Ficedula albicollis, чёрных лебедей, лесной завирушки показана связь вариантов DRD4 с межиндивидуальной вариабельностью дистанций взлёта (Garamszegi et al. 2014; van Dongen et al. 2015; Holtmann et al. 2016, обзор см. Blumstein, 2019). Их укорочение связано с резистентностью особей к общему беспокойству в урбосреде, способностью выделить главное и требующее немедленного реагирования в потоке событий, и сразу отреагировать, переключив поведение без «инерции», обычной для «лесных» птиц.

Городские чибисы на крыше в Манчестере
Отсюда естественна связь DRD4 с урбанизацией видов. Чёрные лебеди вокруг Мельбурна значительно осторожней, чем в городе, их реакция страха и бегства гораздо инерционней, они отлетают дальше и пр. (чем отличаются «городские» особи у большинства видов птиц, Miranda, 2017; Фридман, 2018; Liker, 2020). В городе они подпускали ближе и были устойчивей к беспокойству: эти различия связаны с конкретными генотипами DRD4, различающимися между обоими популяциями (van Dongen et al., 2015). Связь определённых вариантов этого гена с урбанизацией найдена уже у 6 видов птиц (у наполеонова бархатного ткача Euplectes afer — с созданием популяций на новых территориях птицами, улетевшими из неволи, Müller et al., 2014, 2017). Так, в 4-х городских популяциях большой синицы (595 птиц) из Дании в отсеквенированном гене DRD4 найдено 98 однонуклеотидных замен (SNP), ассоциируемых с изменением поискового поведения. Они связаны как с кодирующими, так и с некодирующими участками, первые несут следы позитивного отбора. Наибольшая ассоциация с изменением поисковой активности — с SNP экзона 3 (особенно в 830й позиции), поменьше — с SNP экзонов 2 и 4. Изменения шли параллельно во всех урбопопуляциях, максимум ассоциаций отмечен в гг.Вестерхейде и Ловерсмер. Там же — но не в Boshoek и Wytham Woods — показана связь с исследовательским поведением (Müller et al., 2013b).
Современный город гетерогенен настолько, что там находят место обе альтернативы поискового поведения больших синиц, быстрые vs медленные исследователи, сортируясь между участками урболандшафта с разным характером беспокойства. При урбанизации вида обе стратегии проходят отбор на большую выраженность каждой из них, связанный с изменениями в гене DRD4. Это показано сопоставления данных его секвенирования в городских популяциях с изменениями его SNP при искусственной селекции за или против данных альтернатив. Обе альтернативы поискового поведения больших синиц «находят своё место» в современном городе.
При селекции в пользу «быстрых» или «медленных» исследователей (4 поколения) меняется частота вариантов SNP830T, но не инделя 15, взятого для контроля, т. е. разные аллели SNP830 способствуют проявлению разных типов индивидуальности: С/С — второй, С/Т и Т/Т — первой. Сходные изменения частот данных вариантов отмечены при селекции за и против «быстрого исследования» относительно неселектированной популяции (Fidler et al., 2007).

Молодой сапсан в Бристоле
Отбор «городских» вариантов DRD4 больших синиц сопровождается и эпигенетическими изменениями. В урбопопуляциях уровень метилирования CPG-острова в промоторе и экзоне 3 данного гена (содержит 2 CPG-острова, один из которых заходит в интрон 3) был значимо, но незначительно (на ≈1%) выше как в образцах ДНК из мозга, так и из крови. В городских и в исходных популяциях равным образом CPG-остров в промоторе метилирован слабо (8%) по сравнению с экзоном 3 DRD4 (93%) (Riyahi et al., 2015).
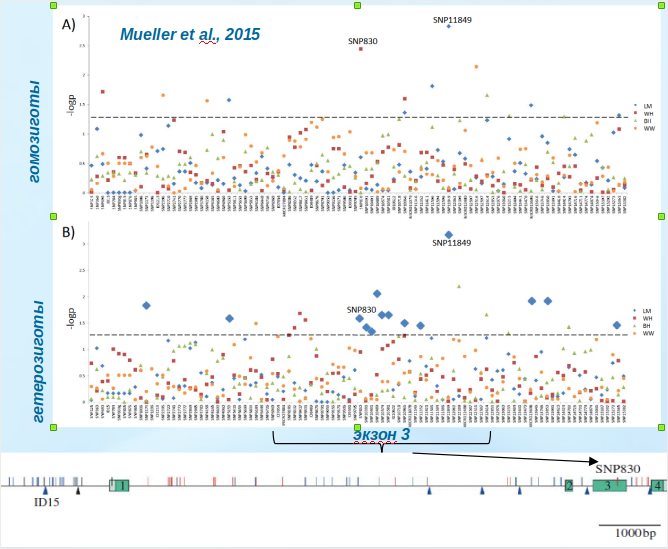
Рисунок 1. Ген DRD4 «городских» больших синиц: CPG-острова и SNP, по которым идёт отбор
Вообще, метилирование ДНК
а) различается между городскими и исходными популяциями, включая локусы DRD4 и SERT;
б) как минимум у больших синиц, земляных вьюрков G.fortis, G.fuliginosa и кроличьих сычей это связано с поведенческими различиями между популяциями;
в) анализ транскриптомов тех же видов показывает устойчивые различия в профилях экспрессии множества разных генов между «городскими» и «исходными» местообитаниями (Riyahi et al. 2015; Verhulst et al. 2016; Watson et al., 2017; McNew et al., 2017).
Что говорит о значимости эпигенетических различий для канализации изменений, связанных с их освоением. Последнее согласуется с важностью любого стресса, особенно в раннем онтогенезе, для программирования изменений, поднимающих устойчивость существования в будущей среде обитания (Suslov, 2013; Суслов, 2014; Роговин, 2019).
Наполеонов бархатный ткач разводился в клетках. Птицы, бежавшие из неволи, создали «городские» популяции в Испании с 1984 г. и в Португалии с 1992 г. В обоих отмечен отбор по гену DRD4 в 16 разных SNP и связь отбираемых SNP с решением проблемных задач в условиях новизны. Отловленных птиц выдерживали в клетках, после чего предъявляли два незнакомых объекта, съедобный и нет. Большинство птиц в этих условиях просто усиливали ненаправленную активность, хотя и не отступали или показывали страх, как «сельские» особи. Меньшинство, напротив, снижало активность и скупо, но целенаправленно исследовало предметы визуально, либо манипулируя с ними. Наибольшая связь с этой более сложной реакций выявлена в Португалии у SNP449 и SNP698, в Испании — у SNP449 (Müller et al., 2014).
У людей DRD4 исследовали как ген урбанизации: выявлено различие по частотам SNP в Мехико и деревенских жителей; в Китае связь не подтвердилась. Частоты DRD4*7R аллелей устойчиво отличаются у детей ненцев, чьи родители живут в городах Ямало-Ненецкого автономного округа vs заняты кочевым оленеводством (исследование проведено в интернатах, Ильинских, Ильинских, 2016). Генотип DRD4*7R — предиктор высокого риска отчисления и плохой успеваемости поведения студентов Сибирского государственного медицинского университета (СибГМУ) и Томского госуниверситета славянского происхождения. На первом курсе из них с таким генотипом было 19,4%, на пятом-шестом — 3,5%, вследствие отчисления по названным причинам. Они же отличались отличались антисоциальным поведением: склонность к выпивке, наркомани и токсикомании, что также стало причиной отчисления. Однако у студентов финно-угорского и монголоидного происхождения происхождения тот же генотип не имеет такого эффекта (Ильинских и др., 2015). Ген-кандидат повышенного внимания (4 работы) и высокой ёмкости рабочей памяти (6), что связывается с «длинным» аллелем: 7 и более повторов в экзоне 3 («короткий» вариант — 2 повтора). Один из факторов развития синдрома гиперактивности-дефицита внимания (17 работ), связываемого с тем же «длинным» аллелем. Связан с агрессией и импульсивностью у людей и прочих млекопитающих (Wush, 2018).
Также привлекает внимание исследователей ассоциации своих полиморфизмов с болезнью Альцгеймера, шизофренией, преэклампсией, восприимчивостью к курению, алкоголю, рисков развития ожирения, посттравматического синдрома, как участник (возможный предиктор) успешности освоения иностранных языков и выявления нюансов своего родного, а также как участник распознавания песен у птиц, связи с восприятием стимулов, оцениваемых как неприятные, как возможный предиктор долголетия, как ген агрессивности, тревожности, сексуальных и ментальных расстройств, обучаемости, адаптации к школе.


Источник The Guardian
Влияние DRD4 на эти поведенческие переменные крайне вариабельно, сильно зависит от социальной принадлежности, языка и культуры испытуемых, а также от способа постановки опыта. В задачах на категоризацию для людей «длинные» аллели DRD4 (7 и более повторов) дают лучшие результаты, чем «короткие». Görlick et al., 2015 выявил различия между людьми с «длинным» и «коротким» аллелем гена в задачах на категоризацию (category learning task) и объём оперативной памяти (OPFAN task).
В первых испытуемые классифицируют объекты, характеризуемые 10 качественными признаками с двумя альтернативными состояниями каждый: голова – 1) ориентация головы (вверх или вперед), 2) челюсти или клюв, 3) форма гребня; нога – 4) толстая или тонкая, 5) пальцы или ласты; цвет – 6) серый или желтый; хвост – 7) тонкий или толстый; тело – 8) форма тела (круг или крест), 9) ориентация полосок на теле (горизонтально или вертикально), 10) полоски на шее – внутри контура шеи или вне контура шеи.
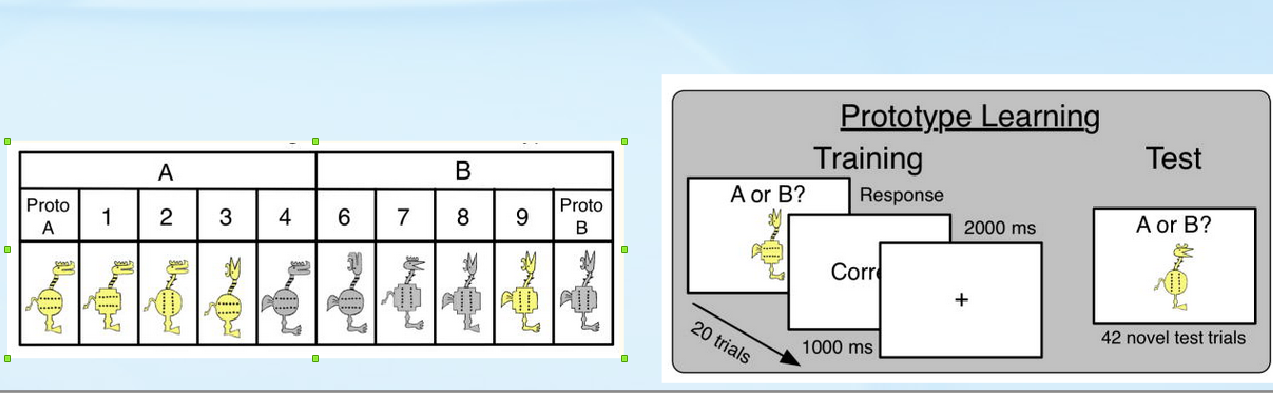
Рисунок 2. Признаки объектов, используемых в задачах на категоризацию, обучение по прототипам и тестирование в работе Görlick et al., 2015.
Участнику предъявляются два прототипа — А и В, у которых все 10 признаков альтернативны. Затем ему задается категория (существенное понятие в биологии) – один из 10 признаков, по альтернативным состояниям которого надо отличать прототипы. Категория выбирается случайно. Например, на левом рисунке один из 4 объектов для любой категории А и В всегда будет лишним и их нужно поменять местами. Его задача: запомнить категорию при предъявлении прототипов, а затем отнести демонстрируемые объекты к той или иной категории. Участник сперва обучается, проходя 20 испытаний по 10 объектов категории A и 10 категории B в случайном порядке. Из них 2 объекта отличались от прототипа по 1 признаку, 3 по 2 признакам, 3 по 3 и 2 по 4 признакам. Неправильная категоризация корректировалась через добавочное время. Потом он проходит сам тест – 42 испытания. Объекты для каждого испытания получают путем изменения у одного и того же прототипа случайно от одного, до 4 признаков.
В задаче на объем оперативной памяти (OSPAN Task) участнику в течение 4000 мс предъявляются карточка с последовательностью из 3-7 букв и арифметическим примером из 4-х действий, затем на 1000 мс — карточка с ответом на пример. Испытуемый, запомнив последовательность букв, должен определить верность решения примера, нажав на кнопку TRUE или FALSE и затем в ходе 2000 мс воспроизвести последовательность букв. Хотя воспроизведение последовательности букв имеет более высокий целевой приоритет, чем решение примера, но верных решений примера должно быть не менее 85% (Conway et al., 2005). Здесь обучения не проводили, сам тест состоял из 15 испытаний по три подхода. Суммарная длина последовательностей букв – 75, последовательности разной длины генерируемых случайно и распределяются по испытаниям нормально.
В одном варианте опыта приняло участие 198 клинически здоровых людей (General), в другом — 187 участников (Screened) успешно прошли международное нейропсихиатрическое интервью (MINI). Носители «длинных» аллелей DRD4 неизменно первенствовали в решении всех когнитивных задач теста. Сравнительно с носителями «коротких» аллелей у них выше а) общее число правильных категоризаций объектов, б) число верных категоризаций схожих (отличающихся 1-2 признаками от прототипа) и в) несхожих (3-4 признаками) объектов (Görlick et al., 2015).
См. данные дисперсионного анализа для а) {ANOVA 2 (screened, general) × 2 (длинные DRD4, короткие DRD4). Длинные DRD4 (M = 0.73, SD = 11) дают лучшую категоризацию, чем короткие DRD4 (M = 0.70, SD = 0.12), F(1, 381) = 4.06, p =0.045, η2= 0.01)}, б) и в) {(ANOVA 2 (screened, general) × 2 (длинные DRD4, короткие DRD4) × 2 (схожие, несхожие). Статус DRD4 F(1, 381) = 4.26, p=0.04, η2 = 0.01}. Длинные DRD4 (M = 0.73, SD = 0.1) дают лучшую категоризацию, чем короткие DRD4 (M = 0.70, SD = 0.12)). Как и ожидалось интуитивно, схожесть с прототипом также вносит вклад в правильность категоризации (F(1, 381) = 3.73, p<0.001, η2 = 0.24). Как и о
Свежие комментарии