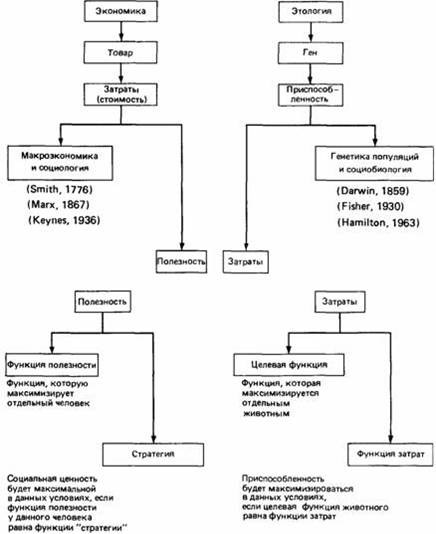
Параллель понятийных аппаратов социобиологии, классического селекционизма и политэкономии: везде критерий выбора поведения из N возможных вариантов — максимизация отношения «платы» и «выигрыша». Проблема в том, что корректное исчисление того и другого требует учёта того, что элементы системы везде не независимы, но связаны специфическими отношениями, и формообразующей роли социальных влияний, «распространяющихся» по этим связям.
Социобиология, увы, этого «не видит» — также как буржуазная политэкономия в противоположность марксистской и эволюционизм Докинза-Гамильтона («отбор генов», управляюющий фенотипами) в противоположность классической СТЭ (отбор фенотипов, под который «подгоняются» изменения на уровне генов, также на на всех прочих). Источник: McFarland, Houston, 1981, цит.по: Д.Мак-Фарленд, 1988. Поведение животных: психобиология, этология и эволюция. Рис.24.15.«Эгоистичный мул»
Содержание
В 2010 г. Александр Кузнецов написал про идеи Докинза «Эгоистичный мул», кольнул, так сказать, в больное (Кузнецов, 2010). Важно подчеркнуть: это именно дружеский шарж, а не испепеляющая сатира (как пытался использовать вопреки предупреждению автора Е.Н.Панов). Описанную ситуацию легко найти в природе: таков кредитогенез у рыб, амфибий и ряда других групп, когда гибридогенный вид в каждом поколении ресинтезируется заново, для чего «эгоистично» эксплуатирует оба родительских. Подобные особи хоть не стерильны, как мулы, но геном одной из форм полностью элиминируется при гаметогенезе, и в следующем поколении для ресинтеза видового фенотипа они должны спариться с одним из родителей (Боркин, 2010, слайды 10, 27–28, также Боркин, Линтвинчук, 2013).
«Эгоистический индивид» обнаруживается лишь в анонимных стаях…
А вот что «убивает» социобиологические концепции напрочь, это примат системности над адаптивностью. Как принцип организации социальной жизни он был выдвинут С.П.Харитоновым (2011) при выделении более и менее предпочитаемых частей колоний. Обнаруживается, что выбор наиболее предпочтительных мест колонии, форм поведения особей в этих местах и/или направлений переселения особей «в эту сторону» внутри колонии часто ведут к понижению репродуктивного успеха (или успеха в конкуренции за территорию, за партнёра, социальный статус на току и т. п. ресурс). А не максимизируют его, как того требуют социобиологические идеализации. Сегодня понятно, что это – универсалия для практически всех видов позвоночных и форм их социальной активности (территориальность, брачное поведение, системы доминирования-подчинения в группах и пр.). Как и для поведения самообеспечения — кормления, использования территории и пр.
 Проявления примата системности над адаптивностью в поведении животных в сообществе подробнее обсуждаются ниже, здесь отметим, что он верен и в «обратную сторону». «Эгоистичные индивиды», соответствующие социобиологическим идеализациям, существуют, но только вне социальной связанности и социальной зависимости, в структурированных сообществах обязательных (Гольцман и др., 1994; Громов, 2008; Фридман, 2013). А именно, в анонимных стаях внегнездового периода, обычных для ряда видов птиц, и в бесструктурных скоплениях особей, привлечённых первичным сигналом (корм, безопасные места ночлега или гнездования и пр.).
Проявления примата системности над адаптивностью в поведении животных в сообществе подробнее обсуждаются ниже, здесь отметим, что он верен и в «обратную сторону». «Эгоистичные индивиды», соответствующие социобиологическим идеализациям, существуют, но только вне социальной связанности и социальной зависимости, в структурированных сообществах обязательных (Гольцман и др., 1994; Громов, 2008; Фридман, 2013). А именно, в анонимных стаях внегнездового периода, обычных для ряда видов птиц, и в бесструктурных скоплениях особей, привлечённых первичным сигналом (корм, безопасные места ночлега или гнездования и пр.).
Так, домовые воробьи Passer domesticus в стае периодически прекращают кормление, поднимают голову и осматривают окрестности, где возможно появления хищника (он нужно обнаружить заблаговременно, чтобы успеть с броском в ближайшее укрытие). Когда они кормятся в плотном скоплении на стационарной кормушке или в рассредоточенной группе на земле, затраты времени на сканирование пространства вокруг в общем падают с ростом размера стаи.
В опытах Elgar et al. (1984) кормовой столик надвое делили перегородкой. Птицы в одной половине не видели собратьев в другой, но могли видеть с присады, откуда слетали к кормушке. Здесь интенсивность осматриваний на кормушке зависела только от числа непосредственно видимых птиц и не зависела от общего числа в группе. Хотя только что, перед слётом, воробей видит всю стаю и при кормлении слышит позывы других птиц за перегородкой, так что мог бы определить их число – но воспринимает лишь своё непосредственное окружение.
В подобных стаях воробьи, овсянки, скворцы и др. виды не координируют моменты осматриваний, хотя таким образом могли бы сильно увеличить время пребывания с опущенной головой (т. е. кормления). У особей, находящихся рядом друг с другом, клювы направлены в разные стороны, притом что неспецифическая стимуляция действует — чем выше среднее число особей, находящихся в поле зрения каждого члена стаи, тем интенсивней он кормится (больше извлекает кормовых объектов в единицу) времени и меньше осматривает при прочих равных. Она же поддерживает высокую синхронность взлётов и точность манёвров стаи, когда индивидов захватывает распространяющаяся волна возбуждения, но, осев на кормление, они вновь делаются независимыми. См. данные по стаям лебедей Cygnus cygnus и C.bewickii (Black, 1988), скворцов Sturnus vulgaris (Fernández-Juricic et al., 2004) и пр.
Ухудшение условий кормодобывания (учащение неудачных зондирований в стае грачей или куликов, большая рассредоточенность семян и других собираемых кормовых объектов) увеличивает частоту и дальность перемещений неуcпешно кормившихся птиц друг относительно друга. Это учащает агрессию и преследования, когда движущиеся особи «натыкаются» на спокойно кормящихся, чем отвлекают уже их и т.д.
Удачно кормящиеся («попавшие» на локальное обилие корма) в такой ситуации интенсифицируют кормодобывание вопреки росту риска хищничества (часто даже в присутствии хищника, как чирки-свистунки Anas crecca в присутствии скопы Pandion haliaetus, Pöysä, 1987). См. данные по серому журавлю Grus grus (Alonso et al., 1986), щеглу Carduelis carduelis (Glück, 1987), грачам Corvus frugilegus (Patterson, 1975). Однако стохастические перемещения «неудачников» побуждают и их учащать осматривания, в конце концов прерывающие кормление. Иными словами, все птицы в стае настолько «эгоистичны», что каждая следует своим собственным стимулам продолжить кормление или прервать его из-за возможной опасности. Поведение соседей здесь фактически не воспринимается до прямого столкновения с ними, исключая кормодобывание.
 Но и они выступают достаточно общим стимулом, интенсифицирующим собственные движения кормодобывания у ближайших соседей пропорционально успешности таковых соседа. Косвенно это влияет на частоту осматриваний, в целом снижая её, но не рождает «коллективной бдительности» — каждая птица следит за соседями и возможным хищником самостоятельно. См. воздействие движений робота на кормящихся мексиканских чечевичников Carpodacus mexicanus (Fernández-Jurisic et al., 2006), или анализ настороженности в стаях юнко Junco hyemalis и воробьиной овсянки Spizella arborea (Lima, 1995).
Но и они выступают достаточно общим стимулом, интенсифицирующим собственные движения кормодобывания у ближайших соседей пропорционально успешности таковых соседа. Косвенно это влияет на частоту осматриваний, в целом снижая её, но не рождает «коллективной бдительности» — каждая птица следит за соседями и возможным хищником самостоятельно. См. воздействие движений робота на кормящихся мексиканских чечевичников Carpodacus mexicanus (Fernández-Jurisic et al., 2006), или анализ настороженности в стаях юнко Junco hyemalis и воробьиной овсянки Spizella arborea (Lima, 1995).
В структурированных сообществах, вроде зимних стай лазоревок Parus caeruleus и больших синиц P.major, ситуация противоположна. Там широко распространены взаимное обучение и «социальная трансляция» инноваций, вроде открывания крышечек из фольги (Aplin et al., 2013, 2015).
Именно из-за «эгоистичности» поведение индивидов во всех этих стаях независимо друг от друга и никак не координируется между собой — хотя это было бы выгодно всем. В силу последнего зависимость уровня риска от его интенсивности и размера стаи колоколообразна: он вновь возрастает при наиболее интенсивном кормлении в крупных стаях. По этой причине особи стараются присоединиться к стае оптимального размера, и лишь голодающие присоединяются к любой. См. данные по желтогорлым овсянкам, Emberiza elegans (Yamaguchi, Yahara, 2002).
Интересно различие в реакции на это взрослых и молодых грачей Corvus frugilegus, кормящихся в зимних стаях. Первые толерантны к рискам учащения агрессивных контактов и выедания корма независимо перемещающимися товарками, они увеличивают темп кормления и достигают большей скорости поступления энергии из пищи (в единицу времени), хотя потребляют более мелких и менее питательных жертв, чем в мелких стаях. Наоборот, молодые грачи не выносят возрастания риска в подобной ситуации — они увеличивают время, затрачиваемое на переходы с места на место и снижают темп кормёжки (Henderson, Hart, 1991).
В обоих случаях оценка риска и/или обилия корма осуществляется локально, по происходящему в 1—1.5 м вокруг индивида, никто не способен осуществить её хотя бы для площади, на которой кормится вся стая. Всецело господствуют близкодействие и «ближний порядок» взаимоотношений индивидов (Coolen et al., 2001).
Из этого исходит Social foraging theory (Giraldeau, Caraco, 2000). Согласно ей, объединение в стаю связано 1) с разделением информации — успешно кормящиеся особи как бы показывают соседям благоприятные микростации и эффективные методы кормодобывания, побуждая осматривать первые и копировать вторые; 2) с возможностью клептопаразитизма части особей («нахлебников», «scroungers») на «честно» добывших корм («producers»), в диапазоне от их сгона, чтобы самому кормиться там же и тем же способом, и до простого изъятия корма (Galef, Giraldeau, 2001). Оба эффекта неизменно локальны (Elgar, 1989; Coolen et al., 2001), дистантное наведение, обслуживающее 1) и 2), а анонимных стаях отсутствует (тем более со специфическими сигналами).
Соответственно, увеличение бдительности с увеличением размера группы (при меньших затратах на осматривания) — индивидуальный, а не коллективный феномен, нивелирующийся возможной дезорганизацией (Roberts, 1996; Bednekoff, Lim, 1998). Так, учащение осматриваний у птиц в стае вызывается интенсификацией перемещений в той же степени, что риск хищничества, и «съедает» вышеописанный выигрыш (Fernández-Jurisic et al., 2004). Интенсификация бдительности при ухудшении видимости (скажем у зябликов Fringilla coelebs, кормящихся семенами), происходит у всех особей одновременно, без попыток «доверия» к бдительности соседа. Также на скорость обнаружения хищника (перепелятника Accipiter nisus) влияет лишь индивидуальная скорость подъёма головы, различия по которой не конвергируют в стае (Whittingham et al., 2004).
Скорее всего, уменьшение риска хищничества в анонимных стаях достигается не за счёт большей (а тем более лучшей) бдительности, а пассивным образом, за счёт массы жертв, которая дезориентирует хищника и сама по себе. Сходная ситуация фиксируется для активной защиты колониальных видов, которая также нескоординирована у разных особей и в целом безуспешна (Толстенков, 2012). В противоположность строгой анизотропности социального пространства колонии, жёстко управляющей территориальным поведением и резидентов, и вселенцев (Kharitonov, 1998; Харитонов, 2011); также как тока и других типов популяционных группировок.
 А интенсификация кормодобывания и сокращение осматриваний в присутствии других членов стаи – частный случай изученного Р.Зайонцем (1965) эффекта социального облегчения. В присутствии “многих глаз” особей, воспринимаемых как “чужие и конкуренты”, как это присуще анонимной стае, у всех её членов растёт общее возбуждение (arousal). Оно улучшает выполнение простых задач (клевки, сканирования, взлёты и повороты все вместе), но ухудшает – сложных (внутреннее структурирование стаи и перераспределение особей в гетерогенной среде). Тогда при первоначальном увеличении числа особей “выигрыш” членов стаи от интенсификации собирания или зондирования превосходит “плату” от дезорганизации перемещений при кормлении и снижения бдительности, но дальше второе начинает преобладать. Что хорошо видно на полевых данных и подтверждается имитационным моделированием (Sirot, 2006).
А интенсификация кормодобывания и сокращение осматриваний в присутствии других членов стаи – частный случай изученного Р.Зайонцем (1965) эффекта социального облегчения. В присутствии “многих глаз” особей, воспринимаемых как “чужие и конкуренты”, как это присуще анонимной стае, у всех её членов растёт общее возбуждение (arousal). Оно улучшает выполнение простых задач (клевки, сканирования, взлёты и повороты все вместе), но ухудшает – сложных (внутреннее структурирование стаи и перераспределение особей в гетерогенной среде). Тогда при первоначальном увеличении числа особей “выигрыш” членов стаи от интенсификации собирания или зондирования превосходит “плату” от дезорганизации перемещений при кормлении и снижения бдительности, но дальше второе начинает преобладать. Что хорошо видно на полевых данных и подтверждается имитационным моделированием (Sirot, 2006).
Неслучайно, что стайный образ жизни сам по себе не коррелирует с большей продолжительностью жизни у исследованных 421 видов североамериканских птиц (Beauchamp, 2010). Точно также размеры переднего мозга у птиц при прочих равных не коррелируют ни со склонностью к стайности во внегнездовое время, ни с размером групп, в противоположность приматам (Beauchamp, Fernández-Juricic, 2004).
…однако немедленно исчезает в социальной жизни
Всё вышеописанное верно также для бесструктурных скоплений индивидов, привлечённых первичным сигналом (локальное изобилие корма, подходящих мест гнездования, безопасных убежищ и пр.) в конкретное место для поселения — чаек, севших на место колонии, тетеревов, собравшихся на току, больших пёстрых дятлов, занимающих одиночные территории в августе-сентябре в плодоносящих сосняках, чтобы охранять их всю зиму и пр. В первые моменты, когда они собрались, но не позже. В подобных скоплениях «эгоистичные индивиды» существуют не дольше свободных радикалов в автокаталитически идущей реакции. Рост локальной плотности особей учащает контакты, те перерастают во взаимодействия с обменом демонстрациями — территориальные, брачные и пр.
По результатам последних животные связываются социальными связями и/или переходят к долговременно-устойчивой эксплуатации территории вместо временных и подвижных участков до этого (т.н. «супертерриториальность», Patterson, 1980). В обоих случаях примат системности над адаптивностью властвует безраздельно, см. ниже.
Одновременно вокализации и/или ритуализованные демонстрации участников взаимодействий (и сам вид победителей и побеждённых), также как трансформированная ими среда обитания (вроде шишек в «кузницах» больших пёстрых дятлов, занявших осенне-зимние территории в обильно плодоносящем сосняке), формируют сигнальное поле группировки. Эти вторичные сигналы привлекают в скопление дополнительных особей – потенциальных вселенцев из «подвижного резерва» популяции (Фридман, Ерёмкин, 2009; Фридман, 2013). Они 1) повышают локальную плотность в формирующейся колонии, на току, в территориальной группировке далеко за пределы экологической ёмкости местообитания, определившей «первичный сигнал» и 2) обостряют конкуренцию за территорию, полового партнёра или социальный статус в группе настолько, что известная часть особей (а в «лучших» поселениях, наиболее притягательных для вселенцев — и большинство) — неминуемо должна проиграть, будучи вытесненной из данного поселения в другие, «похуже», менее плотные и устойчивые из года в год.
Что замыкает положительную обратную связь между плотностью/частотой контактов в бесструктурном скоплении и скоростью/необратимостью его превращения в структурированное сообщество: первое ускоряет второе, а оно, в свою очередь, вздувает первое.
Примат системности над адаптивностью — универсалия социальной жизни
Так исключается сколько-нибудь длительное существование «эгоистических индивидов», как рисует их социобиология, и одновременно участие их в социальной коммуникации — с началом последней животные обращаются в социально связанных и социально зависимых. А их поведение не просто отклоняется от социобиологических идеализаций, но полностью противоположно им, и именно благодаря примату системности над адаптивностью, состоящему в следующем. Индивиды сперва выстраивают видоспецифические социальные связи (в том числе «вкладываются» в поддержание «общей инфраструктуры» информационного обмена в группировке, несмотря на затраты и риск), и только потом конкурируют в их рамках за улучшение статуса («гарантирующего», среди прочего, и больший успех в спариваниях), но лишь при условии устойчивости этих связей.
Так, самцы больших панд не могут «просто спариться» с ближайшей самкой, даже если она доступна и рецептивна, надо сперва «по правилам» посостязаться с соперником (Bian et al., 2013). То же самое видим при образовании пары у домовых воробьёв (Иваницкий, 1994) или воронов (Хейнрих, 1994).
Аналогичный феномен фиксируется у шалашников Chlamydera nuchalis. Здесь самцы привлекают самок, раскладывая «украшения» так, чтобы возникала оптическая иллюзия; чем она выраженней, тем лучше (Endler et al., 2010). Неискусных «иллюзионистов» самки предпочитают меньше, вплоть до полного игнорирования. Однако попытки исследователей им «помочь», перекладывая «украшения» по типу «лучших» особей, ни к чему не привели. В следующие несколько дней самцы всё вернули к прежнему «неудачному» виду, и по-прежнему пытались привлечь им потенциальных подруг (Kelley, Endler, 2012).

Самец большого серого шалашника Chlamydera nuchalis у «шалаша»
То же самое свойственно самцам других видов, спаривающимся на токах: каменным глухарям Tetrao urogalloides (A.B. Андреев, цит. по Харитонов, 2011), турухтанам Phylomachus pugnax (Widemo, 1998), острохвостым тетеревам Centrocercus urophasianus (Patricelli, Krakauer, 2009), шалашникам Ptilonorhynchus violaceus (Paricelli et al., 2002) и пр. В каждый отдельный момент они больше предпочитают демонстрировать на токовой площадке и взаимодействовать с соседями, чем ухаживать за привлечёнными самками – настолько, что им затруднительно «увести» самца с тока и побудить спариваться.
Так, опыты с самками-роботами на току острохвостых тетеревов показывают, что эта стратегия наиболее эффективна в долговременном плане. При каждой данной интенсивности посещений самками тока она обеспечивает устойчивое предпочтение ими «социоцентричных» самцов, наиболее вкладывающихся в выстраивание отношений с соседями, а не в ухаживания – и максимум спариваний за весь период (Patricelli, Krakauer, 2009). С другой стороны, что они же в ухаживании более «отзывчивы» на изменение поведения самки, точней корректируют собственные демонстрации сообразно ответам, а не пробуют «немедленно овладеть», как менее популярные у самок самцы (данные по P.violaceus, Patricelli et al., 2002).
Сюда же относится многажды описанная у колониальных видов верность особей прежним территориям в условиях, когда корма недостаточно для размножения (моевки Rissa tridactyla на Мурмане, Краснов, 1990). Птицы могли бы вести себя «эгоистически» — кто поумнее, переместиться подале, где есть корм, и успешно размножиться, «наплевав» на судьбу группировки. Нет, они «жертвуют» собственным размножением, держат территории в прежней колонии, соседи по-прежнему взаимодействуют между собой в рамках территориальности и взаимного доминирования (Краснов, 1990).
Этим они сохраняют колонию как единицу популяционной системы и мощный источник вторичных сигналов (териологии зовут их сигнальным полем, Щипанов, 2003, 2015), устойчиво привлекающих нерезидентных особей для подселения. Когда временные трудности с кормом кончатся, колония вновь быстро наполнится резидентами. Если бы особи вели себя «эгоистически», им всем вместе пришлось бы искать новые места для образования колоний – а их немного, они требуют уникального сочетания кормовых и защитных условий, так что не факт, что «эгоисты» нашли бы их.

Тем более, при сильном ограничении времени на репродукцию, птицам оказывается выгодней не тратить его на поиски новых мест размножения при каждой отдельной неудаче, но каждый раз «слепо» реагировать на вторичные сигналы и дальше воспроизводить видовые отношения во взаимодействиях друг с другом. В «плохие» сезоны это неэффективно, но «на длинной дистанции» социально-связанные и социально-зависимые особи неизменно выигрывают у «атомизированных» — благодаря пользованию информацией, циркулирующей в социальной системе (т.н. communicative network, Фридман, 2013).
В этих примерах (их можно умножить) поведение особей неизменно преследует долговременный выигрыш, даже если это «невыгодно» с точки зрения максимизации соотношения «платы» и «выигрыша» здесь и сейчас. И даже в экологических, социальных и прочих условиях, когда стремление к долгосрочному выигрышу несовместно с репродуктивным и прочим успехом, обычно оцениваемым этологами как мера приспособленности индивидов. Скажем, люрики Alle alle и большие конюги Aethia cristatella — моногамы; казалось бы, их социальная активность в колонии должна завершаться с образованием пары. Но нет; размножающиеся птицы активно участвуют в «клубах», посещают их на протяжении всего сезона размножения, активно «поют» и вступают в доминантно-субординантные отношения там (Харитонов, 2009).
Здесь опять поддержание социального статуса и активность в «клубах» (т.е. «участие» поведения индивида в поддержании пространственно-этологической структуры колонии) самцам важней собственного размножения. Настолько, что они, уже образовав пару, «поют» в клубах для поддержания и повышения своего статуса, но гонят прочь новых самок, «отказываясь» от возможности спариваний (Харитонов, 2009).
Как показывает С.П.Харитонов (2011, рис.5–6), столь сложная социальная организация не обслуживает размножение индивидов, даже «лучших», скорей отвлекает от него. Люрики и большие конюги размножаются глубоко под камнями, и охраняемые территории на поверхности для размножения значения не имеют. Однако есть и «асоциальные» особи: они не токуют, а прилетая с кормежки, сразу лезут под камни, целиком отдавая себя нуждам размножения (Харитонов, 2011).
То же самое верно для других видов птиц (или млекопитающих) и других социальных систем — тока, территориальности и пр., но полностью противоположно социобиологическим идеализациям, уподобляющим поведение особи расчётливому бизнесмену, действующим всегда и везде на максимизацию «прибыльности» поведения сегодня, в крайнем случае «завтра», но не дальше (Мак-Фарленд, 1988, рис.24.15; Alсock, 1998). Или, точней, движение к долговременному выигрышу возможно здесь лишь как следствие максимизаций серии кратковременных, но не вне этого процесса и тем более не вопреки. Для этого в каждый момент времени выбирается поведение, «наилучшее» по соотношению «платы» и «выигрыша» от его осуществления (из имеющихся альтернатив в виде разных элементов этологического репертуара, альтернативных тактик и стратегий поведения, Maynard Smith, Harper, 2003).
Анизотопность пространства группировок
Действительно, в разных вариантах социальной организации видим, что индивидуальные предпочтения особями разных участков внутри группировки определяются анизотропностью пространства последней (местом в структуре целого), а их проявление часто сопряжено с ущербом для приспособленности особей в долгосрочном аспекте или снижает эффективность поведения здесь сейчас.
Так, большие пёстрые дятлы при занятии одиночных территорий на внегнездовое время неизменно выбирают наиболее плотные поселения, хотя там занять их и удержать наиболее затруднительно из-за максимально ожесточённой конкуренции резидентов, отчего высока доля «неудачников» в первом или втором. Вторгаясь на чужие участки и обороняя собственные от вторжений соседей, они всю зиму стараются «переделить в свою пользу» общее пространство поселения. Помимо соседей, владельцы территорий находятся «под давлением» активных вселенцев, пробующих закрепиться, заняв участок территории в наименее охраняемой области – на краю группировки или на стыках территорий. И затем расширять его до обычных размеров с одновременной интенсификацией охраны, рекламирования пространства и патрулирования границ (Фридман, 1995а, 1998).
Неуспешность охраны территории (учащение поражений, снижающих вероятность победы в n последних стачках ниже некого минимума, и следующее за ними сокращение её площади ниже предела в 2000–2100 м2) вызывает её потерю. «Неудачники», не сумевшие закрепиться в поселении в первые дни после прибытия, или удержать территорию после более продолжительного периода охраны, составляют 60–95% птиц, привлечённых в плотные поселения в конце лета, и 15–45% – занявших участки и удерживавших их >3-5 дней, выстроивших там ночёвочное дупло. При прочих равных (общая численность вида в данной местности, урожай семян в этом году и пр.) доля «неудачников» положительно зависит от плотности, рост которой увеличивает риск «неудачи» и немедленно, и потом (Фридман, 1995а, 1998).
Причём второй ниже, но равномерен в течение всей осени и зимы, и не снижается по мере «знакомства» птиц в группировке и с установлением персонализованных отношений между ними. Подавляющая часть этих особей перемещается в ближайшие (примерно треть) и более дальние (две трети) поселения с меньшей плотностью и межгодовой устойчивостью населения. Они тем менее привлекательны для первопоселенцев, чем ниже их плотность в прошлые годы, в связи с чем их местоположение и состав населения сильнее варьируют по годам (Фридман, 1995а, 2003).

Сигнальное поведение большого пёстрого дятла (по: Blume, 1963,1967; Симкин, 1976):
1 — «раздвоение хвоста» и демонстрация рулевых (поза агрессии и возбуждения), 2 — задержанное нападение и угроза, 3 — защитное развёртывание крыльев, поза страха, предупреждения, запугивания, против дуплового конкурента, а также в первые дни тока в связи с так называемым «преследованием», 4 — поза «дремоты» в перерывах борьбы, 5 — встреча будущих брачных партнёров, 6 — угроза птенцу при распадении выводка, 7 — борьба двух самок у ночёвочного дупла, справа в смешанной позе «дремоты» и «противостояния» — кочующая птица, 8 — поза «противостояния».
Чем ниже плотность, тем более группировка пополнится «неудачниками», вытесненными из более привлекательных поселений данной местности, и чем позже там занимает участки большинство резидентов, держащихся там всю зиму. Срок занятия территорий в разные годы сильно варьирует с середины июля и до начала сентября: он зависит как от интенсивности размножения вида в данном году, интенсивности позднелетней дисперсии, так и урожая семян. Но относительный «нуль отсчёта» в каждом сезоне – позднелетнее занятие первых территорий в наиболее плотных поселениях прошлых лет (Фридман, 1995а, 2003).
Отсюда следует первый вопрос. Почему значительная (иногда большая) часть популяции идёт на крупные дополнительные затраты, «делая крюк» в первоначальных попытках закрепиться в самых плотных поселениях, при высокой вероятности вытеснения, потери времени и бессмысленности произведённых усилий? Можно ведь было сразу прибыть в более разреженные поселения, где занятия территории «по плечу»? Хуже того, пребывание в нетерриториальном статусе, долгие перемещения между поселениями практически у всех видов птиц – и дятлов, и гаичек, и других видов синиц – крайне рискованны для особей (обзор см. Фридман, 2013). Однако естественный отбор не только не элиминировал этот риск, но и сделал его нормативным; это не только не объяснимо социобиологическими теориями, но и противоположно им.
Вторая проблема того же рода – заняв территорию, дятел предпочтительно расширяет её именно к центру поселения, а не к периферии, хотя в первом случае сопротивление выше, а вероятность успеха – ниже. В территориальной активности больших пёстрых дятлов вторжения на участки соседей с последующей маркировкой/патрулированием новых границ криком «кик» (в сентябре и марте также барабанной дробью[1]) чередуются с охраной «ядра» собственной территории от вторжений соседей. С последующей маркировкой их центров и прежних границ, если их «удалось отстоять», при неудаче новые границы маркирует только захватчик, хозяин лишь центр участка (Фридман, 1995а, 1998).
Чем большая доля конфликтов выигрывалась владельцем в прежние 3–5 дней сравнительно с соседями, тем чаще он «демонстрирует амбиции» – вторгается к ним сам в момент, когда сопротивляться им труднее всего, они сами проиграли конфликт с третьей особью. Или когда сосед долбит шишку, что заставляет прерваться и после конфликта лететь за новой, тем самым снижая эффективность кормодобывания. Для того и другого птицы непрерывно отслеживают перемещения и взаимодействия соседей, чему помогает рекламирование территории криком «кик» (временной профиль волны «киканий» меняется в зависимости от того, победил дятел в недавнем конфликте или проиграл, и насколько «убедительно[2]», Фридман, 1998). И наоборот, «неудачные» относительно соседей дятлы преимущественно защищаются от попыток захватов, совершаемых в основном в те же «неудобные» для них моменты (Фридман, 1998).
Все попытки расширения охраняемого пространства идут в первую очередь в сторону центра поселения: вторжения происходят преимущественно через сектора периферии участка, обращённые в сторону центра поселения, защищаются же наоборот, больше границы, «обращённые к периферии» последнего. В такой ситуации в центре (функциональном, не обязательно геометрическом) уже через 1–2 месяца охраны участков, к концу сентября–началу ноября, автоматически оказываются особи с наибольшей успешностью охраны пространства. Прочие просто не удержатся под постоянным «давлением» вторжений более периферийных особей или соседних – на «обращённые к центру поселения» части их собственных территорий.
Фактически складывается анозотропность пространства группировки. Его наиболее центральные участки максимально привлекательны для захватчиков, т.е. выше по «качеству» с точки зрения всех резидентов, включая самых периферийных и неуспешных (Фридман, 1995а, 1998).
Подобная ситуация сохраняется всю зиму, пока в ходе весеннего возбуждения резиденты не станут всё чаще вместо охраны пространства сближаться с соседями противоположного пола и пробовать образовывать пары. Успех в этом процессе заканчивается объединением прежних одиночных территорий, поиском места для гнезда и пр., событиями, знаменующими прекращение внегнездовой территориальности и характерной для неё «связи всех со всеми» внутри поселения. Брачные пары сперва охраняют объединённую территорию от появившихся по соседству пар и одиночек, однако всё больше оказываются независимы от них.
Собственно территориальная агрессия здесь слабеет, но усиливается – в контексте «натравливания» самками своих самцов на соседей и конкуренции холостых самцов, зимовавших в другом месте, и недавно занявших территории, за внимание самки. Хотя птицы их продолжают охранять примерно до начала яйцекладки, такой социальной связанности и социальной зависимости соседних владельцев, как в осенне-зимний период, с момента образования пары уже не наблюдается[3] (Фридман, 1995б, 1998).
Так или иначе, устойчивость градиента «качества» территорий при сохранении центростремительности территориальных амбиций всех особей, даже периферийных, противоположно требуемой социобиологами максимизации краткосрочного выигрыша. «Материальная выгода» резидентов состоит в удержании и, возможно, расширении территории, позволяющей устойчиво кормиться всю зиму с наименьшими затратами времени и энергии, и минимальным риском её потерять. Но дятлы, и самые «неуспешные», жертвуют «выгодой» ради «качества» – положения относительно центра. Расширяя охраняемое пространство в эту сторону, они резко увеличивают риск территорию потерять, так как сталкиваются с владельцами, охраняющими территорию ещё более эффективно чем они и их соседи «сбоку».
Однотипная анизотропность обнаружена на токах и в колониях — видах внутрипопуляционных группировок с многажды большей плотностью, напряжённостью контактов особей и, соответственно, интенсивностью конкуренции за территорию, партнёра и/или социальный статус. Там также выделяются наиболее предпочитаемая область («центр, в функциональном, а не геометрическом смысле), «субцентр» – прилегающие к нему области и «край».
У колониальных видов описаны те же центростремительные направления территориальных амбиций, что у больших пёстрых дятлов в одиночно-территориальных группировках и на токах тетеревов Lyrurustetrix (Лысенко, 1987). Помимо попыток расширить территории в сторону центра, за время существования колонии озёрные чайки Larus ridibundus переселяются, «перенося» территории в эту сторону. Лишь единицы гнездятся в той части колонии, где они заняли первую в данном сезоне территорию, большая часть пар её «переносит» и всегда — в более предпочитаемые всеми районы группировки.
Особенно это заметно, когда колония L.ridibundus включает в себя несколько сплавин с центром на одной из них. По оценкам С.П.Харитонова (2011), за сезон от 30 до 71% чаек с разных сплавин пробуют переселиться в более предпочитаемые части колонии, реально переселяется 12–36% птиц. Помимо переселения в центр, есть примерно в 2,5 раза меньший поток переселений в обратную сторону, сходный с «оттоком» неудачников в охране территорий у больших пёстрых дятлов.
Самый важный момент: вопреки социобиологическим представлениям переселения не ведут к росту репродуктивного успеха (разному на разных сплавинах и т. д. частях колонии), но во многих ситуациях снижают его. Так, в колонии озёрных чаек на оз.Киёво в 1983 г.начал разрушаться большой плавающий остров, бывший центром колонии, из-за чего каждый сезон разорялись тысячи гнезд. В эти же годы успех размножения на береговых сплавинах, где таких разорений не было, был намного выше. Однако же чайки по-прежнему переселялись в центр, но не на береговые сплавины (Харитонов, 2011, рис.4).
Хуже того, на фоне падения численности колонии в 1983–87 гг. в центре «освободились вакансии», он стал доступней для чаек с краю колонии, что дополнительно увеличило переселение туда, при игнорировании стабильных, успешных мест размножения на береговых сплавинах. Последствия для размножения индивидов и колонии в целом были печальны — её площадь стала сокращаться от периферии к центру до полного исчезновения (Харитонов, 2011).
Сходное поведение демонстрируют и другие колониальные виды птиц, также как виды, гнездящиеся «под охраной колоний», вроде черношейных поганок Podiceps nigricollis. Они предпочитают гнездиться в центре колонии озерных чаек, – там, где они их беспокоят больше всего. Возникает парадоксальная ситуация, когда кладка поганок ниже в месте тяготения большинства пар (Чухарева, Харитонов, 2009). В одной из изученных С.П.Харитоновым локальных популяций малых крачек Sterna albifrons более половины особей (по ряду признаков – наиболее “качественных”) из года в год упорно гнездятся там, где постоянно возникающие нагонных ветры неизменно губят все гнёзда. У тонкоклювых кайр Uria aagle в Тихоокеанском секторе Арктики при меньшей стабильности гнездовых биотопов на птичьих базарах есть наиболее предпочитаемые карнизы, куда стремится существенно больше птиц, чем во все прочие. В подобной колонии бывает много незанятых карнизов, но ищущие место кайры предпочитают уже заселенные (Харитонов, 2011, рис.3).

Эти примеры показывают, что озерным чайкам (тонкоклювым кайрам, малым крачкам, и пр.) «психологический комфорт» от «движения» в наиболее предпочтительный центр колонии важнее успеха размножения, максимизация которого требовала бы «двигаться» в противоположную сторону. В столь странной с точки зрения естественного отбора привязанности (повторяющейся у разных видов, не только колониальных) естественно видеть «проекцию» на индивидуальном уровне примата системности над адаптивностью. Последний был выдвинут С.П.Харитоновым (2011) при выделении более и менее предпочитаемых частей колоний, когда обнаружилось, что выбор наиболее предпочтительных мест колонии, форм поведения особей в этих местах и/или направлений переселения особей «в эту сторону» внутри колонии часто ведут к понижению репродуктивного успеха (или успеха в конкуренции за территорию, за партнёра, социальный статус на току и т. п. ресурс), а не максимизируют его, как требуют социобиологические идеализации.
По-видимому, примат системности над адаптивностью – универсалия социальной жизни позвоночных вообще, включая использование территории и перемещения особей, составляющих «подвижный резерв» популяции. Что согласуется с выводом С.П.Харитонова об отсутствии принципиальной разницы между колонией и надколониальной системой: предпочтения особями разных частей внутри первой и переселения в более предпочтительные части организованы также, как между отдельными колониями во второй (Kharitonov, 1998; Харитонов, 2011).
Из общих представлений социобиологов следуют частные концепции. Они описывают структуру разных видов активности как оптимальную для особи по главному критерию максимизации краткосрочного выигрыша (т.е. отношения «benefit» к «cost» при каждом следующем выборе форм поведения). C их точки зрения все ныне существующие формы прошли интенсивный отбор на способность вести себя именно так, а иные стратегии были отброшены как эволюционно нестабильные (Maynard Smith, Harper, 2003; Searcy, Novicky, 2005). Для кормодобывания и долговременного использования территории индивидами (т. н. пространственно-ориентированное поведение) это концепция оптимальной фуражировки (optimal foraging theory, Krebs, Davies, 1997); для расселения за пределы местностей, где находятся территории – гипотеза приспособленности резидентов (resident fitness hypothesis, Anderson, 1989), для социальных взаимодействий между компаньонами, включающих организованный обмен демонстрациями между компаньонами, т. е. коммуникацию — концепция манипуляции (Фридман, 2013, т. 2, табл.3).
Использование пространства: долгосрочная устойчивость важней «оптимальности фуражировки»
Однако реальность опрокидывает все эти теории. Во всех перечисленных животные неизменно «менее оптимальны», чем постулируют социобиологи. Их поведение организовано так, что обязательно включает формы активности, расходы на которые «здесь и сейчас» представляются лишними и ненужными с точки зрения максимизации краткосрочного выигрыша (и эти расходы довольно значительны от 5–10% до 15–25% бюджета времени и энергии). Однако они совершенно необходимы с точки зрения долгосрочной устойчивости развёртывания данной формы активности особи в реальной среде, с характерным для неё уровнем «шума» и «помех» для этой последней в виде изменений доступности корма, погодных условий, персонального состава партнёров, их выбытия от разных причин, непредсказуемого для особи и неконтролируемого ею[4].
Так, использование территории птицами или млекопитающими неоптимально, ибо всегда включает дальние исследовательские выходы, за пределы индивидуального участка — пространства, ресурсы которого эксплуатируются изо дня в день. Следуй поведение особей социобиологическим концепциям, выходы были б элиминированы как «плата», не приносящая «выигрыша» непосредственно в данный момент, как в условиях рынка на Северном флоте «сэкономили» на спасательных средствах, не требующихся каждый день — пока не случилась авария подлодки «Курск»…
Так, территориальные самцы певчих воробьиных «обычно» охраняют территорию, поют и кормятся в пределах охраняемого пространства, но «время от времени» совершают дальние выходы и рейды по окружающим территориям, а часто и в соседнее поселение. Это «время от времени» составляет у разных видов от 10 до 40% светлого времени, но так как дальние выходы совершаются периодически, выплесками, неподготовленный наблюдатель просто считает перерыв в наблюдениях из-за потери резидента, и лишь радиопрослеживание с массовым цветным мечением изменило картину (Иваницкий, 1998).
Вне своих территорий самцы (реже самки) почти не поют, территориальной активности не проявляют, поэтому могут кормиться, (почти) не навлекая агрессии владельцев. Большая часть рейдов осуществляется внешне бесцельно и вроде безрезультатно, но на деле периодические рейды позволяют «улавливать» нерегулярно появляющиеся возможности обоим полам увеличить репродуктивный успех (которая обычно ещё и выпадает с не очень большой вероятностью). Например, вступить во внебрачные копуляции с резидентными особями или занять вторую территорию и петь там вдали от основного участка, чтобы привлечь туда самок, оставшихся холостыми (Akҫay, Rougharden, 2007).
Дуплогнёздники во время “выходов” инспектируют дупла соседних пар своего и других видов, в т.ч. во время гнездого периода, отрывая время от насиживания и выыкармливания. Так, у мухоловки-пеструшки Ficedulahypoleuca на Звенигородской биостанции МГУ за 15 сезонов зафиксировано 760 визитов, 94% из них сделано самцами, холостыми и размножающимися, 5% — самками, 1% — молодыми в ювенильном пере. В основном птицы проявляли исследовательскую активности, иногда и кормили чужих птенцов (Ильина, 2012).

Мухоловки-пеструшки у гнезда
Уже концепция биологического сигнального поля Н.П.Наумова (1973, цит. по Щипанов, 2000) заставляет предполагать, что перемещения особей в пространстве популяции не хаотические, а в известной степени канализированы (а тем более переселения индивидов внутри группировки или в ближайшие соседние). Сейчас проясняются конкретные факторы канализации.
Во-первых, это предпочтение переселяющимися особями среды, уже изменённой деятельностью конспецификов: большие Rhombomys opimus и монгольские песчанки Meriones unguiculatus охотней заселяют пустующие «колонии» своего вида, чем не обжитые территории. Поселившись, при выборе участка обитания зверьки предпочитают микростации со следами жизнедеятельности других особей, и в особенности запаховыми метками (Громов, 2000).
Наличие определённых путей перемещения показано, в частности, для лесных полёвок и лесной мыши Apodemus sylvaticus. В ряде экосистем старые тропинки мелких млекуопитающих хорошо просматриваются на протяжении нескольких лет. При наступлении подъёма численности первые зверьки ловятся именно на тропинках. Специальные эксперименты показывают структурированность перемещений особей даже в однородной среде. На площадке в 1 га исследовали посещение нор малого суслика с использованием полосок закопченной бумаги. Малые суслики обитали на участке, лишённом растительности, при равномерном покрытии территории норами (190 вертикальных и горизонтальных входов). Полоски бумаги вводили на ночь в отверстие нор. Оказалось неожиданно, что полоски заслежены проходящими через площадку домовыми мышами. Число заслеженных (мышами) полосок в текущие сутки коррелировало с числом встреченных на площадке нерезидентных мышей (r=0,75). Распределение мест регистрации мышиных следов в пространстве показало перемещение зверьков между сусличьими норами по двум определённым маршрутам (Щипанов, 2000).
Эти пути «намечаются» во время регулярных выходов особей за пределы собственной территории. Индивидуальное мечение мелких млекопитающих показало, что у видов с постоянными участками обитания их владельцы совершают исследовательские кормовые выходы на расстояние до 0,5-1 км от основной территории зверька. Особенно типичны они для полёвок рода Microtus и Gletrionomys: эта оборотная сторона территориального поведения выполняет здесь целый ряд функций на уровне популяции (Бененсон, Садыков, 1989). Выходы равномерны во всех направлениях, хотя чаще идёт вдоль «экологических коридоров» и направляющих линий в рельефе местности, существенных для зверьков.
Постоянное совершение выходов снабжает резидентов информацией о состоянии среды и качестве местообитаний, о доступности корма в пределах территории, намного превышающей их собственный участок. Она «востребуется» резидентом а) при невозможности обитания на прежнем участке, б) при расселении его потомства (в обоих случаях временные поселения возникают именно в местах исследовательских кормовых выходов, а исходная плотность поселившихся зверьков пропорциональна частоте посещения).
Живущие на Иремеле (горный массив на южном Урале) красные полёвки G.rutilis, постоянно обитающие в наиболее благоприятных участках со стабильной и высокой численностью («резервациях»), ежедневно посещают окружающие стации независимо от наличия условий для формирования временных поселений. Среднее удаление от основного участка составляет 300-500 м в день. Расселение аналогично первой фазе исследовательского выхода, лишь в один из них зверёк не возвращается, а остаётся на новом участке. Соответственно, минимальная группировка, динамика которой тождественна или изоморфна популяционной — это население резерваций и всех производных от неё временных поселений в зонах сезонного и транзитного режима использования (Бененсон, Садыков, 1989; Щипанов, 2000).
Наконец, только выходы дают возможность резиденту на своём небольшом участке сохранять оптимальную структуру рациона, поддерживать оптимальный баланс количества и качества потребляемого корма в условиях постоянных изменений растительных сообществ, продуктивности и доступности разных видов кормов, включая изменения, вызванные потреблением самого зверька и сверхпотреблением проходящих особей или заходящих соседей. Чтоб поддерживать оптимальный баланс, рацион должен постоянно меняться, сохраняя постоянство некоторых пропорций, скажем между основными кормами, дающими нужную сумму калорий, и «деликатесами», дающими белки, витамины, и другие ценные компоненты в необходимом количестве.
Поэтому конфигурация индивидуального участка под воздействием изменений растительности должна своевременно и адекватно меняться вслед за изменениями среды, особенно представляющими собой долговременные тренды. То же относится и к концентрации убежищ. В условиях варьирующего риска нападения хищника территория и способ её использования зверьком, также как и отношения между зверьками в сообществе должны быстро и адекватно меняться, чтобы не возрастал уровень риска, а вокруг активного животного всегда нашлось несколько убежищ на дистанции мгновенного броска в укрытие (Смирин, 1984).
Исследовательские кормовые выходы позволяют резиденту решить эту задачу прогнозирования главных тенденций изменения экологической среды. Благодаря им животное «исходит» из кормовых и защитных условий много более обширного пространства, чем его собственный участок обитания, что даёт возможность более точного сравнения качества стаций последнего в более репрезентативном ряду местных биотопов. Естественное следствие – лучшие разделение времени в суточном бюджете активности между посещением разных парцелл растительности на собственном участке и способность индивидуального пространства активности адекватно «рассыпаться» на функциональные зоны, постоянно видоизменяться во времени и пространстве в соответствии с главными тенденциями средовых изменений.
Благодаря постоянным изменениям конфигурации и функциональной структуры территории внутри неё всегда находится та мозаика растительных сообществ, которая может обеспечить устойчивость эксплуатации необходимого объёма кормовых и защитных ресурсов. Эта адекватность и опережающий характер реагирования невозможны без постоянных исследовательских выходов на некоторую минимальную дистанцию. Во всяком случае, жёсткое ограничение выходов в виде ограждения пространства вокруг зверька, соответствующего средней площади индивидуального участка, ведут к быстрой гибели от голода при истощении предпочитаемых кормов и явном недоиспользовании субоптимальных и второстепенных (Шварц, 2004), тогда как в природе зверьки вполне выдерживают эту пропорцию. Этот же результат получен при ограждении кормящихся выводков куликов-песочников, что говорит в пользу универсальности подобного механизма «улавливания» тенденции средовых изменений дальними выходами.
Для всех видов песчанок характерны спонтанные выходы оседлых зверьков-резидентов за пределы участка обитания, иногда на значительное расстояние, с последующим возвращением. «Выходящие» зверьки активно исследуют запаховые метки и следы жизнедеятельности соседних особей, вступают с ними в разнообразные социальные контакты. У «несоциальных» видов песчанок спонтанные выходы совершают в основном взрослые самцы, вообще более активные и имеющие большее индивидуальное пространство по сравнению с самками (Громов, 2000).
У «социальных» монгольских песчанок такие «рейды» одинаково часто совершают и взрослые особи обоего пола, и подросшие сеголетки, особенно из первых, весенних, выводков. Но целью спонтанных выходов у монгольских песчанок чаще всего является территория соседних социальных групп: «разведывательные рейды» нарушают границы семейно-групповых участков. Исследовательская активность с дальними выходами за пределы территории особенно усиливается в годы депрессии, а также в ситуации резкого снижения численности популяции (дератизационные обработки). Следовательно, у «несоциальных» видов соответствующая активность служит «социальному отталкиванию» и стимулируется необходимостью поиска свободных ресурсов (территория, партнёр) для данного индивида. То же самое поведение у «социальных» видов обслуживает «социальное притяжение» между особями в популяции, и поэтому управляется чисто социальными стимулами (Громов, 2000, 2008).
Периодические рейды резидентов по чужим территориям быстро завершаются реальным переселением в «лучшую позицию» в другой или в той же самой группировке, приобретением второй территории (политерриториальность) или, например, внебрачными копуляциями.
У краснокрылого трупиала Agelaius phoeniceus взрослые особи (уже размножавшиеся в данной группировке) всегда предпринимают перемещения в соседние при возможности увеличить успех размножения (пытаясь занять там территорию, чаще посещаемую самками). Достоверно увеличивается репродуктивный успех самцов, переместившихся на дистанцию <200 м. Он постоянен у самцов, сохранивших прежнюю территорию, но снижается у переместившихся больше чем на 200 м (снижение пропорционально дальности, Beletski, Orians, 1987, цит.по Фридман, Ерёмкин, 2009).
В большинстве колоний A.phoeniceus со стабильными условиями существования возможность увеличить репродуктивный успех за счёт переселения возникает нечасто. Она почти всегда связана с событиями внутри группировки — разделом освободившихся территорий между соседями или их расширением в результате экспансии самца вслед за самкой, подыскивающей место для гнезда за пределами первоначального участка. Потому большая часть самцов очень привязана к своим территориям, и направленные переселения между колониями совершает незначительная – но всегда существенная – часть группировки. Это так называемые «подвижные резиденты» как особая категория индивидов, которую следует отличать от нетерриториальных птиц «подвижного резерва» популяции (Picman, 1987; Dickinson, Lein, 1987, цит. по Фридман, Ерёмкин, 2009).

Шалфейная овсянка
Другой пример: у шалфейной овсянки Amphispiza belli при перемещении качественные характеристики территории не меняются но, как правило, увеличивается площадь участка, от которой зависит репродуктивный успех владельца. Средняя дальность перемещения отрицательно коррелирует с успехом размножения в предшествующем сезоне. Перемещения территорий регистрируются на протяжении всей жизни; после каждого перемещения репродуктивный успех территориального самца не сильно, но значимо увеличивался, зато прогрессивно сокращалась средняя дальность перемещений (Petersen, Best, 1987, цит.по Фридман, Ерёмкин, 2009).
Направленные переселения резидентных особей столь же свойственны территориальным воробьиным, даже несмотря на меньшую продолжительность жизни. Особь может совершать переселения на всём протяжении жизни, они всегда направлены из разреженных и неустойчивых поселений в ближайшие к ним более оптимальные. Например, в многочисленных популяциях молодых оттесняют в пессимальные местообитания, и затем на протяжении жизни они постепенно “возвращаются” в плотные поселения в лучших местообитаниях. Неуспех таких переселений «по восходящей» обычно побуждает особь оставить местную популяцию, принять участие в более дальней дисперсии – вплоть до перемещений типа выселений или инвазий (Иваницкий, 1998).
Такие переселения зафиксированы у мухоловки-пеструшки (Высоцкий, 1991, цит.по Фридман, Ерёмкин, 2009) и даже у пеночки-веснички Phylloscopus trochilus. У веснички раньше прилетающие молодые занимают лучшие территории в оптимальных биотопах, откуда их вытесняют старые птицы в пессимальные местообитания. Обретая опыт успешного размножения, птицы в следующие сезоны постепенно возвращаются на место первоначального выбора. Из-за высокой ежегодной смертности на прежние территории возвращается 20-50% популяции, из них направленные переселения предпринимает 18% особей. Средняя дальность переселений взрослых резидентов до 100-300 м, молодых – до 1300 м (Jakobsson, 1987; Rädesater et al., 1991, цит.по Фридман, Ерёмкин, 2009).
Как и в социальной активности, в использовании территории социальная связанность индивидов даже не «пересиливает» постулируемую социобиологами максимизацию «прибыльности» поведения каждого, а просто «не считается» с ней. Точнее, последнее не появляется там и тогда, где этому противоречит структура социальных связей животного, выступающих для него источником психологического комфорта. Они снижают неопределённость среды до оптимального уровня, повышая предсказуемость событий в ближайшем окружении особи, почему «связанные» особи в ограниченном держатся вместе и перемещаются координированно (данные по большим синицам, Ильина и др., 2009). Атомизированные же в анонимных стаях – перемещаются независимо и поддерживают индивидуальную дистанцию, неизменно отвечая агрессией даже на случайное нарушение.
Поэтому связанность облегчает участникам таких отношений исследовать незнакомые пространства и новые ситуации, вместо обычной для птиц в анонимной стае неофобии. Так, у больших синиц исследовали социального фактора в освоении новой среды: простой (тест «открытое поле») и сложной (вольеры 2х2х2 м с присадой, кормушкой с кормом и убежищем). На 15 мин. в 24 вольеры выпустили одиночных синиц, а в 49 – по две разнополые особи одновременно, фиксируя число обследованных объектов, латентный период обследования каждого объекта (период времени от начала сеанса до первого посещения объекта), а также период времени до начала кормежки. При одиночном обследовании индивидуальность особи чётко просматривалась: синицы, более подвижные в «отрытом поле» быстрее обследовали объекты в вольере. В присутствии партнёра скорость освоения вольер не была связана с индивидуальной подвижностью в «открытом поле», а скорость перемещений снижалась, уступая исследовательской активности (Ильина, 2010).
В опытах Firth et al. (2015) со свободноживущими парами больших синиц создавали конфликт между кормлением и социальной связанностью, выставляя на занятом ими участке леса автоматические кормушки, доступные только одной особи (открываются по радиомаячку на конкретной особи). Кормушки для половины пар настраивали так, что самца и самку вынуждали кормиться максимально далеко друг от друга, в противоположных частях участка, другая половина была контролем, кормушки партнёров находились поблизости. Все брачные партнёры шли на заметное сокращение поступления пищи, чтобы держаться рядом друг с другом и сопряжённо перемещаться по участку (что резко расходится с предсказаниями теории «оптимальной фуражировки»). У части из них наблюдали «делёжку» корма, добытого одной из птиц, с партнёром, т. е. для поддержания социальной связанности часть птиц может в большей или меньшей степени превращаться из «добытчиков» («producers») в «нахлебников» («scrounders») (Firth et al., 2015).
Тот же самый эффект проявляется при переселении между группировками и занятии территориальных «вакансий» там особями из «подвижного резерва» популяции (подтверждая вывод С.П.Харитонова об отсутствии принципиальных различий в организации подобного поведения внутри популяционных группировок «между ними»). У всех видов мелких млекопитающих, где наблюдается быстрое восстановление за счёт направления (сигнальным полем) потоков нерезидентов в место локальной катастрофы (виды с хорошим развитием т. н. восстанавливающей подсистемы, «тип 2» по классификации Н.А.Щипанова), это происходит за счёт того, что эмигранты не только прибывают физически в нужное место, но и размножаются интенсивней, чем в стабильных поселениях (эти особи образуют т. н. контролирующую подсистему внутри популяции (Щипанов, 2003, 2016). Последнее происходит за счёт того, что среди вселенцев присутствуют взрослые самки, часть которых находится на последних стадиях беременности (Getz et al., 2005). С т.з. индивидуальной приспособленности это выглядит бессмысленным, и даже вредным.
Если бы поведение особей соответствовало социобиологическим идеализациям, самки, тем более беременные, демонстрировали бы наибольшую оседлость, так как и эффективность кормления, и безопасность (величина, обратная риску гибели от нападения хищника) у мелких млекопитающих на каждой территории определяются знанием участка (Anderson, 1989). Из этого исходит гипотеза межполовых различий в дисперсии Пауля Дж. Гринвуда: активности во время натальной дисперсии самцы превосходят самок, когда они конкурируют непосредственно за них, а самка выбирает самца. Если же самка выбирает территорию, и самцы конкурируют за лучшие территории, а не напрямую за самок, то всё наоборот (Greenwood, 1983).
Она была проверена Чарльзом А. Брандтом на одном из видов-исключений — пищухе Ochotona princeps. Оказалось, что здесь репродуктивный успех самок определяется исключительно типом биотопа и не лимитируется «качеством» самца. Репродуктивный успех самцов же коррелирует с привлекательностью занятого участка для самки, а не с их собственным «качеством». Соответственно, и молодые самки расселяются на большие расстояния, чем самцы, что согласуется с гипотезой Гринвуда И то же самое обнаруживается у другого вида-исключения – летучей мыши Saccopteryx bilineata (Brandt, 1985).

Американская пищуха, пика Ochotona princeps
Однако в обоих случаях беременные самки должны быть максимально привязаны к наименьшему пространству, известному им наилучшим образом; если они и участвуют в переселениях, то молодыми, т. к. индивидуальный отбор поддерживает оседлость самок пропорционально их репродуктивному вкладу (Anderson, 1989). Также предполагается асимметрия в соотношении полов среди переселенцев, но не репрезентативность по отношению к популяции исхода.
Вне популяционных регуляторов это вполне справедливо, см. выше; но, под действием сигнального поля и др. механизмов, направляющих потоки переселенцев так, что восстановление оказывается быстрым и точным (по уровню численности), особи в популяции более управляемы системным целым, чем собственными «интересами». Как минимум у 4-х видов серых полёвок отмечено несмещённое расселение, когда выборка нерезидентов не отличается от демографического состава популяции (вопреки гипотезе Гринвуда). У малой белозубки состав нерезидентного населения определяется случайной последовательностью просыпания зверьков. Смещение соотношения полов в сторону самок у «восстанавливающих» нерезидентов также описано у обыкновенной бурозубки Sorex araneus и пр. (Щипанов, 2016).
Следовательно, гипотеза Гринвуда относится лишь к переселениям между устойчивыми группировками, резиденты в которых эффективно поддерживают определённый уровень численности вила в данной местности, но не могут интенсифицировать размножение даже в благоприятных условиях, из-за развитой территориальности и других способов исключения из репродукции части взрослых особей. Но не к восстановлению после истребления (Шилова, 1993; Щипанов, 2003, 2016).
Обсуждение
Из вышеприведённых данных следует, что достичь цели, представляющейся социобиологам главной (максимизировать итоговую приспособленность) поведение животных «может» только после того, как перемещения и взаимодействия их создали и стали поддерживать видоспецифический паттерн социальных связей, в их рамках и при условии их устойчивости, а не самому по себе. Структуры отношений, в рамках которых возможны «состязания» за итоговую приспособленность (они для социобиологов альфа и омега эволюции поведения), даны индивидам извне, и относятся к надындивидуальному уровню организации.
Так, у 19 видов певчих воробьиных численность в каждом из сезонов размножения отрицательно коррелирует с плохими погодными условиями в апреле-начале мая, то есть в период распределения территорий, формирования пространственно-этологической структуры местных популяций, но не коррелирует с погодными условиями в июне-июле, непосредственно в период выкармливания птенцов. Важно подчеркнуть, что в работе рассматриваются весьма протяжённые временные ряды наблюдений, охватывающие около 60 лет, и включающие в себя всё разнообразие «хороших», «плохих» и «промежуточных» сезонов размножения в количестве, достаточном для репрезентативности и статистического анализа (Соколов, 1999). Иными словами, особи «вкладываются» в поддержание пространственно-этологической структуры группировок в апреле – мае, а «выигрыш» получаю в июле-августе, причём он зависит от обстоятельств первого, а не второго.

Двулинейный мешкокрыл Saccopteryx bilineata
Как бы ни были особи возбуждены, активны и «эгоистичны», им не дано эти отношения «пересмотреть» и направленно «изменить в свою пользу», можно только «сломать», выйти за их пределы. Но за это немедленно следует «наказание» исключением из социальных связей, и прости-прощай итоговая приспособленность. Хотя видоспецифичный паттерн социальных связей идеален (это «реальность, лишённая телесности»), для особей он не меньшая, а большая реальность, чем непосредственно хищники, пища, территориальные оппоненты и брачные партнёры «своего» вида.
Ведь появление этих ценных (для максимизации приспособленности) объектов и развитие взаимодействия с ними воспринимается особью не непосредственно, но через изменение социальных связей, «в перекрестье» которых развивается индивидуальность животного. Так паук обнаруживает жертву и партнёршу по дрожанию нитей паутины, и также оценивает их, выбирая что делать. См. данные о том, что стрессом можно «заразиться», он распространяется по социальным связям, у зебровых амадин Taeniopygia guttata (Monaghan et al., 2011) и людей (Engert et al., 2014). Причём во втором случае личные отношения для «заражения» необязательны, достаточно наблюдения, что хорошо соответствует большей социальности нашего вида.
Никогда не происходит иначе: если максимизация размножаемости входит в противоречие с поддержанием социальных связей, особи неизменно выбирают второе, даже при риске вообще не размножиться. Причём данное поведение наиболее характерно именно для видов с максимальной напряжённостью конкуренции за партнёра — обладающих развитыми «брачными украшениями» и/или спаривающимися на токах. Хотя во всех перечисленных случаях птицам достаточно «сделать шаг в сторону» — перейти в поселение с большей ресурсообеспеченностью, с лучшими перспективами привлечения самок для данной особи, здесь неуспешной, и их приспособленность выросла бы с нуля до значимой величины. Но нет, они «верны» уже образованным социальным связям, и могут наращивать приспособленность «лишь в их рамках». По той же причине не распадаются уже образованные брачные связи у действительно моногамных видов, даже если это смешанная пара разных видов с плохими перспективами размножения, или однополая пара, образованная из-за нехватки самок — и даже если потом появляются активно токующие партнёры «своего» вида.
Таким образом, главный прокол социобиологии следующий. Социальная связанность и социальная зависимость индивидов целиком и полностью ликвидирует «эгоистичность». Особи начинают действовать строго в пользу долговременного выигрыша, даже когда это сопряжено с ущербом витальным интересам или дарвиновской приспособленности самих индивидов.
Есть ли здесь исключения? Да Исключения — т. н. «несоциальные» виды позвоночных (норвежский лемминг Lemmus lemmus, азиатский бурундук Eutamias sibiricus,песчанки полуденная M.meridianus и тамарисковая M.tamariscinus), составляющие сугубое меньшинство в «своих собственных» родах и семействах (Громов, 2000, 2008). Они противопоставляются «социальному» большинству видов по следующим признакам.
1. В особо продуктивных биотопах «несоциальные» виды достигают не меньшей плотности, чем социальные, как полуденная песчанка на сенобазах. Но это не увеличивает частоту контактов — зверьки дифференцируют время и место активности так, чтобы по-прежнему не встречаться, и сталкиваются только случайно. Последнее сопровождается неритуализованной агрессией без последействия, долговременно меняющего поведение победителя и проигравшего в сторону большей комплементарности поведения и повторения взаимодействий, как у «социальных» видов, т. е. к связыванию регулярными отношениями (Гольцман и др., 1994).
2. У «социальных видов» грызунов структура связей в сообществе, основанная на отношениях агрессивного доминирования, настолько устойчива, что не меняется от уменьшения/увеличения агрессивности индивидов фармакологическим действием антидепрессантов/нейролептиков (Громов, 2000).
Ещё более интересные опыты того же рода были проведены С.А.Шиловой (1993) в природе с разными видами грызунов, различающихся сложностью социальной системы – и соответственно, степенью дифференциации поведенческих ролей у особей разного социального статуса, развитостью ритуализованной агрессии во взаимодействиях, степенью социальной асимметрии, устанавливаемой в ходе каждого социального контакта и её последействием – влиянием на поведение обоих особей в последующих взаимодействиях и пр. Это монгольская пищуха Ochotona pallasi, длиннохвостый суслик Spermophilus undulatus, полуденная песчанка, лесная и домовая мышь (эти 2 вида – в вольере), монгольская песчанка. Виды ранжированы по возрастанию степени социальности и усложнению социальной системы. Им давали галоперидол, резерпин, амизил и пр. (Шилова, 1993: 144–153).
Вполне предсказуемо выяснилось, что снижение агрессивности у видов с наиболее просто устроенной социальной системой (суслик, пищуха) разрушало её полностью или в существенной степени, у видов со сложной – не влияло или почти не влияло. В этом эволюционная выгода от увеличения сложности надындивидуальных систем, объединяющих «эгоистических индивидов» в одно целое, сопрягающих их активностью и делающих её из исходно независимой – комплементарной в рамках антисимметричных социальных ролей (доминант – подчинённый, самец – самка в паре и пр.). Чем сложней социальная структура группировок, чем выше «степень социальности» вида, тем выше устойчивость к «возмущениям», связанным с изменением состояния индивидов, их численности, персонального состава, да и экологическим изменениям вокруг тоже. А это означает всё большую автономизацию этих особей от внешней, экологической среды, при увеличении регулярности, предсказуемости среды социальной (Шилова, 1993: 144–153).
У несоциальных видов эти системные эффекты отсутствуют, у «промежуточных», вроде использованных С.А.Шиловой — ослаблены в сравнении с наиболее «социальными».
3. У «социальных» видов плотностно-зависимые регуляторы численности популяции вызываются к действию её разрежением в не меньшей степени, чем переуплотнением (т. е. они максимально подвержены принципу Олли, это т. н. сильный регуляционизм в популяционной динамике). У «несоциальных» они максимально ослаблены или отсутствуют вовсе, как у норвежского лемминга. Там регуляция происходит «по-мальтусовски» — неизбирательным истреблением и/или голодом после вспышки массового размножения (стохастицизм в популяционной динамике). У «промежуточных» видов данные регуляторы «запускаются» преимущественно переуплотнением (слабый регуляционизм).
Фактически особи здесь — альтруистические машины, заточенные эволюцией на соединение друг с другом в соообщества, сообществ — в группировки, чтобы координированно (контрастируя с прежним «эгоизмом») создать и воспроизводить в этой системе поселений социальную организацию вида, её характерный паттерн пространственно-этологической структуры. Лишь после того как активность животных «выстроила» соответствующую структура связей, в её рамках поддерживаются инфраструктуры, обеспечивающие определённого рода долговременный выигрыш, всем участникам, «вложившимся» в поддержание всего перечисленного — коммуникативная сеть группировок, их сигнальное поле в виде среды, изменённой активностью особей и пр. — только тогда эти последние действовать на максимизацию собственной приспособленности, и лишь в рамках созданной ранее структуры связей.
Отсюда примат системности над адаптивностью представляется а) несовместным с социобиологическими концепциями, б) реальной универсалией социальной жизни позвоночных, вследствие чего первые не проходят фальсификационный тест. Тогда конкуренция за партнёра оказывается лишь «техническим средством» укрепления наиболее важных связей в социальной организации вида (через «напряжение» их конкурентной борьбой или «отдачей» от неё, с последующим корректированием селективными процессами). Иногда эти связи лежат в области брачного поведения, чаще нет; это от вида зависит.
Данный вывод сполна относится к социальному поведению людей в обществе (с поправкой, что межвидовым различиям социальной у животных изоморфны различия общественного устройства разных стран и народов, Фридман, 2016). Действительно, примат системности над адаптивностью зафиксируется в известных нейроэкономических экспериментах по Ultimatum Game, когда люди стабильно отвергают возможность «получить хоть что-то» и любое разделение ценностей, отличающееся от равного. Это, можно сказать, общечеловеческая константа — соответствующие результаты стабильно получены на всём разнообразии обществ от охотников-собирателей до западных клерков. Первобытные люди несколько чаще отвергали предложения получить сильно больше половины, видимо потому что в их обществе «власть рождается из отдачи, подчинение из получения» (пословица африканского народа кпелле). Ситуация не менялась даже в случаях, когда речь шла о суммах, сравнимых с месячным заработком игроков (Боулс, Гинтис, 2006).
Во всех этих случаях принятие или отвержение испытуемыми выгодного, но подлого предложения с резко неравным разделением средств, отклонявшееся от поровну в обе стороны более чем на 20%, определяется активностью двух зон мозга. Это островковая кора (anterior insula), вовлечённая в обработку негативной эмоциональной информации и особенно активна при эмоции отвращения; её активность была пропорциональна степени несправедливости предложения. С другой стороны, это верхние области лобной коры (dorsolateral prefrontal cortex, DLPFC) правого полушария и поясная извилина (anterior cingulate cortex), активирующиеся при запуске когнитивных процессов самоконтроля и внутренних конфликтах. Причём по соотношению активности островковой коры и DLPFC можно предсказать, будет полученное игроком предложение принято (вторая активируется много сильнее первой) или отвергнуто (наоборот) (Ключарёв и др., 2011).
Эти данные можно интерпретировать двояко. Первый вариант — от животных мы унаследовали «дикую» тягу к равенству и справедливости, которая проявляется в эмоции отвращения к их рациональной противоположности — сделке подлой, но «лучше чем ничего». Но, по счастью, она контролируется и преодолевается «чисто человеческим», в виде контроля со стороны лобной коры. Авторы обзора явно симпатизируют данной точке зрения, почему называют несправедливый выбор «рациональным», хотя он, понятное дело, нерационален для принявших его в долгосрочном аспекте. Ведь с учётом долговременных перспектив всем участникам выгодней именно равенство в разделении ресурсов – по тем же причинам, что привязанность к территориям на току, в колонии и пр. в условиях средовой и демографической стохастичности.
Вторая интерпретация противоположна. «Животное» в нас — это как раз калькуляция «платы» и «выигрышей», побуждающее принять несправедливое предложение, что уподобляет людей участникам анонимных стай, также максимизирующих соотношение «выигрыша» к «плате» здесь и сейчас. «Движение» к долгосрочному выигрышу и долгосрочной устойчивости здесь невозможно. Однако социальные связи этот животный эгоизм в прямом смысле слова трансформируют и преодолевают, заставляя предпочитать долговременный выигрыш (постоянное соблюдение равенства во всём круге участников) краткосрочному (принять «рациональное» предложение сейчас, а там хоть трава не расти). В этом случае контроль лобной коры (нашего «разума») связан именно с тем, чтобы сохранить устремление к долгосрочному выигрышу, не купиться за предложение, опасное в перспективе, сохранять примат системности над адаптивностью, как компас сохраняет норд. Недавно произведённые красивые опыты показывают, что верна именно вторая модель.
Когда при помощи транскраниальной магнитной стимуляции (ТМС) временно подавляли активность DLPFC в правом полушарии, принятие испытуемыми несправедливых предложений в игре «Ультиматум» гораздо чаще, чем при подавлении активности DLPFC в левом полушарии. Иными словами, после инактивации DLPFC испытуемые активнее начинают демонстрировать утилитарное поведение, принимая невыгодные или несправедливые предложения. Однако это не влияло на субъективную оценку справедливости/несправедливости предложений: испытуемые с инактивацией правой DLPFC, отмечали, что «их возмущала несправедливость ситуации, но отвергнуть несправедливые предложения после ТМС-стимуляции они не могли» (Ключарёв и др., 2011).
Полученный результат хорошо согласуется с тем, что у животных индивиды, интегрированные социальными связями, перестают быть «эгоистичными» и действуют строго в пользу долговременного выигрыша даже супротив собственных витальных интересов или дарвиновской приспособленности. Или с тем, что у людей в экономических играх, воспроизводящих устойчивое природопользование, именно «коллективисты голосуют за будущее» (Hauser et al., 2014).
Заключение
Вышепроведённый анализ показывает, что самый убийственный аргумент против социобиологии – особи разных видов позвоночных отселектированы всей предшествующей эволюцией так, что ведут себя как эгоистические индивиды лишь в бесструктурных скоплениях. А как только окажутся в «структурированных» сообществах, так немедля «общественное оказывается выше личного» — социальное поведение «отшлифовано эволюцией» так, что точность выстраивания видоспецифических отношений на уровне всего сообщества каждому индивиду важней роста собственной приспособленности независимо от других. Соответственно удовлетворение витальных потребностей и прочие действия, увеличивающие приспособленность, начинаются только после выстраивания видоспецифических социальных отношений и при условии его успешности; ради первого особь легко жертвует вторым, но не наоборот. Это проявляется что в территориальных конфликтах и ином социальном поведении, что в эксплуатации территории.
Причин этому две: 1) приспособленность нельзя максимизировать в одиночку, надо вступить в специальные отношения с прочими, чтобы (при успехе конкурентной борьбы) эксплуатировать их как социальный ресурс, а для этого отношения следует подчинять видовой норме, чем точнее, тем лучше. 2) Самому их менять как минимум неразумно, ибо силы и знания индивидов ограничены, куда лучше воспользоваться информацией, распространяемой по коммуникационной сети, сигнальному полю и другим «инфраструктурам», поддерживающим «дальний порядок» внутри популяции и информирующим индивидов о важных событиях на временах и пространствах, много больше их непосредственного окружения. А для этого надо подчиняться правилам сети организации взаимодействий, частью которой становится активность индивидов.
Библиографический список
Бененсон И.Е., Садыков О.Ф. Концепция и модель популяционной динамики мелких млекопитающих в гетерогенной среде/ И.Е.Бененсон, О.Ф.Садыков// Журн. общей биологии. — 1989. — Т.50. — №2. — С.244—259.
Боркин Л.Я. Сетчатое видообразование — эволюция не по Дарвину? [Электронный ресурс] / Л.Я. Боркин// 2010. — Режим доступа: http://www.bio.spbu.ru/science/conference/darwin/materials/Borkin.pdf. Дата обращения: 6.12.2016.
Боркин Л.Я., Линвинчук С.Н. Гибридизация, видообразование и систематика животных / //Труды ЗИН РАН . — 2013. — Т.317. Приложение № 2. — С.83—139 .
Боулс С., Гинтис Г. Вальрасианская экономическая теория в ретроспективе/ С. Боулс, Г.Гинтис// Истоки: из опыта изучения экономики как структуры и процесса. — М.: ГУ ВШЭ, 2006. — С.301—337.
Глушенков О.В. О недейственности активных форм коллективной защиты колониальных птиц/ О.В. Глушенков// Теоретические аспекты колониальности у птиц. — Ростов н/Д: изд-во ЮНЦ РАН, 2012. — С.57—69.
Гольцман М.Е., Попов С.В., Чабовский А.В. и др. Синдром социальности: сравнительное исследование поведения песчанок/ М.Е. Гольцман, С.В. Попов, А.В. Чабовский и др // Журн. общей биологии. — 1994. — Т.55. — №1. — С.49—69.
Громов В.С. Пространственно-этологическая структура популяций грызунов/ В.С.Громов. — М.: Товарищество научных изданий КМК, 2008. — 581 с.
Иваницкий В.В. Воробьи и родственные им группы зерноядных птиц: поведение, экология, эволюция/ В.В.Иваницкий. — М.: KMK Scientific Press Ltd, 1998. — 147 с.
Иваницкий В.В. Пространственно-ориентированное поведение птиц/ В.В. Иваницкий // Орнитология. — Вып. 28. — М.: Изд-во МГУ, 1998. — С. 3—23.
Ильина Т.А. Роль социального фактора в освоении новой среды у большой синицы (Parus major) в условиях ограниченного пространства/ Т.А.Ильина// Зоол. Журн. — 2010. — Т.89. — №9. — С.1131—1138.
Ильина Т.А. Феномен визитёрства у мухоловки-пеструшки (Ficedula hypoleuca Pall., Passeriformes, Aves) в гнездовой период/ Т.А.Ильина// Вестник Моск. ун-та. Сер. 16: Биология. – 2012. – №2. – С.49–53.
Ильина Т.А., Керимов А.Б., Иванкина Е.В. Предпочитаемая дистанция как показатель социальных взаимоотношений больших синиц (Parus major) в условиях дефицита пространства/ Т.А.Ильина, А.Б.Керимов, Е.В.Иванкина // Зоол. ж. — 2009. — Т. 88. — №.1. — С..71—77.
Ключарёв В.А., Шмидс А., Шестакова А.Н. Нейроэкономика: нейробиология принятия решений/ В.А.Ключарёв, А.Шмидс, А.Н.Шестакова // Экспериментальная психология. — 2011. — Т.4. — № 2. С.14–35.
Краснов Ю.В. Социальная организация чайковых птиц в условиях многолетнего кризиса кормовой базы/Ю.В.Краснов// Современные проблемы изучения колониальности у птиц: Материалы 2 совещания по теоретическим аспектам колониальности у птиц, Мелитополь, 26 сент.—1 окт., 1988. Симферополь, Мелитополь, 1990. — С. 77—80.
Кузнецов А. Эгоистичный мул/ А.Н. Кузнецов// Химия и Жизнь. — 2010. — №4. — С. 28—31.
Лысенко И.Г. Пространственно-этологическая структура тетеревиного тока/ И.Г.Лысенко// Биологические основы охраны и воспроизводства охотничьих ресурсов. Сб. научных трудов. — М.: ЦНИЛ Главохоты РСФСР, 1987. — С.14—25.
Мак-Фарленд Д. Поведение животных. Психобиология, этология и эволюция. / Д. Мак-Фраленд. — М.: Мир, 1988. — 520 с.
Смирин Ю.М. Влияние фактора опасности на характер взаимоотношений зверьков в искусственных группировках рыжих полёвок (Gletrionomysglareolus) и лесных мышей (Apodemussylvaticus)/ Ю.М.Смирин // Зоол.ж. — 1984. Т.LXIII. — №6. — С.888—898.
Соколов Л.В. Популяционная динамика воробьиных птиц/ Л.В. Соколов// Зоол. Журн. — 1999. — Т. 78. — № 3. — С. 311—324.
Фридман В.С. О территориальном поведении большого пёстрого дятла в поселениях высокой плотности: типы социальной структуры в осенне-зимний период и их смена/ В.С. Фридман // Орнитология. — Вып.26. — М.: изд-во МГУ, 1995а. — С.3—16.
Фридман В.С. Брачное доминирование у пестрых дятлов (роды Dendrocopos и Picoides) и его причинный анализ/ В.С. Фридман// Бюлл. МОИП. Сер.биол. — 1995б. — Т.100. — №5. — С.44—61.
Фридман В.С. Социальная структура популяций Dendrocopos major в изменчивой среде: как сохранить единство при разнонаправленных адаптациях особей?/ В.С. Фридман// Жизнь популяций в гетерогенной среде. Материалы II Всеросс. Семинара. — Йошкар-Ола, 1998. — Кн.1. — С.267—284.
Фридман В.С. Особенности территориального поведения больших пестрых дятлов в московских и подмосковных лесах: запрещающие и предписывающие факторы освоения городской среды/ В.С. Фридман // Животные в городе. — Мат-лы II научно-практ. конф. М.: РоСХА, 2003. — С.151—155.
Фридман В.С. От стимула к символу: сигналы в коммуникации позвоночных / В.С. Фридман. — в 2-х тт. — М.: изд-во URSS, 2013. — 560 с. и 424 с.
Фридман В.С. Социальная эволюция Homo sapiens: взгляд зоолога/ В.С. Фридман// Социально-экологические технологии. — 2016. — №1. — С.76—88.
Фридман В.С., Ерёмкин Г.С. Урбанизация «диких» видов птиц в контексте эволюции урболандшафта/ В.С.Фридман, Г.С.Ерёмкин. — М.: изд-во УРСС, 2009. — 240 с.
Харитонов С.П. Структура колонии и поведение высокосоциальных морских птиц – люриков (Alle alle) и больших конюг (Aethia criststella)/ С.П. Харитонов // Экология, эволюция и систематика животных (Материалы Всерос. науч.-практ. конф. с междунар. участием 17–19 ноября 2009 г.) — Рязань: Голос губернии, 2009. — С. 21–22.
Харитонов С.П. Пространственно-этологическая структура колоний околоводных птиц/// С.П.Харитонов// Зоол.журн. — 2011. — Т.90. — №7. — С.846—860.
Хейнрих Б. Ворон зимой /Б.Хейнрих. — М.: изд-во «Мир», 1994. — 321 с.
Чухарева И.П., Харитонов С.П. О пространственной структуре поливидовых колоний, основанных чайковыми птицами, на водоемах Южного Зауралья/ Чухарёва И.П., Харитонов С.П. // Поволжский экол. Журн. — 2009. — № 3. — С. 238–248.
Шварц Е.А. Сохранение биоразнообразия: сообщества и экосистемы/ Е.А.Шварц. — М.: Товарищество научных изданий КМК, 2004. — 112 с.
Шилова С.А. Популяционная экология как основа контроля численности мелких млекопитающих/ С.А. Шилова. — М.: Наука, 1993.— 203 с.
Щипанов Н.А. Некоторые аспекты популяционной устойчивости мелких млекопитающих/ Н.А. Щипанов // Успехи совр. биол. — 2000. — Т.120. — №1. — С.73—87.
Щипанов Н.А. Популяция как единица существования вида. Мелкие млекопитающие/ Н.А. Щипанов// Зоол.ж. — 2003. — Т.82. — №4. — С.450—469.
Щипанов Н.А. Функциональная структура популяции и видовое разнообразие. Мелкие млекопитающие/ Н.А. Щипанов // Сборник трудов Зоологического музея МГУ им. М.В. Ломоносова. — 2016. — Т.54. — С.478—513.
Akçay E., Roughgarden J. Extra-pair paternity in birds: review of the genetic benefits/ E.Akçay, J. Roughgarden// Evolutionary Ecology Research. – 2007. – V.9. – P.855–868.
Alonso J. A., Veiga J. P., Alonso J.C. Time budgeting and social structure of Common Cranes Grus grus wintering in Iberia/ J. Alonso, J.P. Veiga, J.C. Alonso //Ricerche di Biologia della Selvaggina, Bologna, 1986. — V.10 – С.1—14.
Alcock J. Animal Behavior: An Evolutionary Approach/ J. Alcock. — Sunderland: Sinauer, 1998. — 6th ed. — 550 pp.
Anderson P.K. Dispersal in rodents: A resident fitness hypothesis. / P.K. Anderson. — Special publication. — № 9. —American Society of Mammologist, 1989. — 141 p.
Aplin L.M., Sheldon B.C., Morand-Ferron J. Milk bottles revisited: social learning and individual variation in the blue tit, Cyanistes caeruleus/ L.M. Aplin, B.C. Sheldon, J. Morand-Ferron/ Anim. Behav. — 2013. — V.85. — №6. — Р.1225—1232.
Aplin L.M., Farine D.R., Morand-Ferron J., Cockburn A., Thornton A., Sheldon B.C. Experimentally induced innovations lead to persistent culture via conformity in wild birds / L.M. Aplin, D.R. Farine, J. Morand-Ferron, A.Cockburn, A.Thornton, B.C. Sheldon // Nature. — 2015. — V.518. — №7540. — P.538—541.
Beauchamp G. Group-foraging is not associated with longevity in North American birds / G. Beauchamp// Biol. Lett. — 2010. — V.6. — P.42–44.
Beauchamp G., Fernández-Juricic E. Is there a relationship between forebrain size and group size in birds? / G. Beauchamp, E. Fernández-Juricic //Evolut. Ecol. Res. — 2004. —V.6. — P.833–842.
Bednekoff P.A., Lim S.L. Randomness, chaos and confusion in the study of antipredator or vigilance/ P.A. Bednekoff, S.L. Lim // Trends Ecol. and Evol. — 1998. — №7. — Т.13. — С.284—287.
Bian X., Liu D., Zeng H., Zhang G., Wei R., Hou R. Exposure to Odors of Rivals Enhances Sexual Motivation in Male Giant Pandas [Электронный ресурс]/ X. Bian, D. Liu, H. Zeng, G. Zhang, R. Wei, R. Hou. // PLoS ONE. — 2013. — V.8. — №8. — Режим доступа: https://www.academia.edu/31821827/%D0%A7%D1%82%D0%BE_%D0%BD%D0%B5_%D1%82%D0%B0%D0%BA_%D1%81_%D1%81%D0%BE%D1%86%D0%B8%D0%BE%D0%B1%D0%B8%D0%BE%D0%BB%D0%BE%D0%B3%D0%B8%D0%B5%D0%B9_2016_http:/dx.doi.org/10.1371/journal.pone.0069889http://dx.doi.org/10.1371/journal.pone.0069889. Дата обращения: 20.10.2016.
Black J. M. Preflight signalling in swans: a mechanism for group cohesion and flock formation/ J.M. Black //Ethology. – 1988. – Т. 79. – №. 2. – С. 143—157.
Brandt C.A. The evolution of sexual differences in natal dispersal: the test of Greenwood’s hypothesis// Migration: mechanisms and adaptive significance/ C.A. Brandt// Conrtribution in Marine Science, Suppl. (Univ. of Texas). — 1985. — V.27. — Ed. M.A.Rankin. — P.386—396.
Coolen I., Giraldeau L.-A., la Voie M. Head position as an indicator of producer and scrounger tactics in a ground-feeding bird/ I. Coolen, L.-A. Giraldeau, M. la Voie // Anim. Behav. — 2001. — V.61. — P.895—903.
Elgar M., Burren P., Posen M. Vigilance and perception of flock size in foraging house sparrows (Passer domesticus L)/ M. Elgar, P. Burren, M. Rosen// Behaviour. — 1984. — V.90. — №4. — Р.215—223.
Elgar M.A. Predator vigilance and group size in mammals and birds: a critical review of the empirical evidence/ M.A. Elgar// Biol. Rev. — 1989. —V. 64. P. 13—33.
Endler J.A., Endler L.C., Doerr N.R. Great bowerbirds create theaters with forced perspective when seen by their audience/ J.A. Endler, L.C. Endler, N.R. Doerr // Current Biology. — 2010. — V.20. — P. 1679–1684.
Engert V., Plessow F., Miller R., Kirschbaum C., Singer T. Cortisol increase in empathic stress is modulated by emotional closeness and observation modality/ V. Engert, F. Plessow, R. Miller, C. Kirschbaum, T. Singer // Psychoneuroendocrinology. – 2014. — V.45. – №7. – P.192–201.
Fernández-Juricic E., Siller S., Kacelnik A. Flock density, social foraging, and scanning: An experiment with starlings / E. Fernández-Juricic, S. Siller, A. Kacelnik// Behav. Ecol. — 2004. — V.15. — №3. — Р.371—379.
Fernández-Juricic E., Gilak N., McDonald J.C., Pithia P., Valcarcel A. A dynamic method to study the transmission of social foraging information in flocks using robots/ E. Fernández-Juricic, N. Gilak J.C. McDonald, P. Pithia, A. Valcarcel// Animal Behav. — 2006. — V.71. — P. 901—911.
Firth J.A., Voelkl B., Farine D.R., Sheldon B.C. Experimental Evidence that Social Relationships Determine Individual Foraging Behavior/ J.A. Firth, B. Voelkl, D.R. Farine, B.C. Sheldon// Curr. Biol. — 2015. — V.25. — №23. — Р.3138–3143.
Galef B.G. Jr, Giraldeau L.-A. Social influences on foraging in vertebrates: causal mechanisms and adaptive functions/ B.G. Galef, Jr, L.-A. Giraldeau// Anim. Behav. — 2001. — V.61. — №1. — Р.3—15.
Getz L.L., McGuire B., Carter C.S. Social organization and mating system of free-living prairie voles, Microtus ochrogaster: a review/ L.L. Getz, B. McGuire, C.S. Carter// Acta Zoologica Sinica. — 2005. — V.51. — №2. — 178–186.
Giraldeau L.-A., Caraco T. Social Foraging Theory/ L.A.Giraldeau, T. Caraco. — Monographs in Behaviour and Ecology. — Princeton, New Jersey: Princeton University Press, 2000. — 376 pp.
Glück E. Benefits and costs of social foraging and optimal flock size in goldfinches (Carduelis carduelis)/ E.Glück// Ethology. — 1987. — V.74. — P.65—79.
Greenwood P.J. Mating systems, philopatry and dispersal in birds and mammals/ P.J. Greenwood // Anim.Behav. — V.28. — №4. — Р.1140–1162.
Hauser O. P., Rand D. G., Peysakhovich А., Nowak М.А. Cooperating with the future/ O. P. Hauser, D. G. Rand, A. Peysakhovich, M. A. Nowak.// Nature. – 2014. – V. 511. – P. 220–222.
Henderson I.G., Hart P.J. Age-specific differences in the winter foraging strategies of Rooks/I. G. Henderson, P.J. Hart// Oecologia. — 1991. — V.85. — P.492—497.
Maynard Smith J., Harper D. Animal Signals: Oxford Series in Ecology and Evolution/ J. Maynard Smith, D. Harper. — NY: Oxf. Univ. Press, 2003. — 166 pp.
Monaghan P., Heidinger B.J., d‘Alba L., Evans N.P., Spencer K.A. For better or worse: reduced adult lifespan following early-life stress is transmitted to breeding partners/ P. Monaghan, B.J. Heidinger, L. d’Alba, N.P. Evans, K. A. Spencer // Proc. R. Soc. B. – 2011. – DOI: 10.1098/rspb.2011.1291.
Kelley L.A., Endler J.A. Male great bowerbirds create forced perspective illusions with consistently different individual quality/ L.A. Kelley, J.A. Endler // PNAS. — 2012. — V.109. — №51. — Р. 20980—20985.
Kharitonov S.P. Waterbird colony structure: system approach/ S.P. Kharitonov// Ornithologia. — V.28. — Moscow: MSU Publ, 1998. — P.26—37.
Krebs J.R., Davies N.B. (eds.). Behavioural Ecology: An Evolutionary Approach. / J.R. Krebs, N.B. Davies. — 4rd edition. — Oxford, Blackwell, 1997. — 464 pp.
Lima S.L. Back to the basics of anti-predatory vigilance: the group-size effect/ S.L. Lima// Anim. Behav. — 1995. — V.49. — P.11—20.
Patricelli G.L., Krakauer A. Measuring male Greater Sage-Grouse response to female signals during courship: an experiment with a robotic female [Электронный ресурс]/ G. Patricelli, A. Krakauer // Behav. Ecol. — 2009. — Режим доступа:doi:10.1093/beheco/arp155. Дата обращения: 15.09.2016.
Patricelli G.L., Uy J.A.C., Walsh G., Borgia G. Male displays adjusted to female’s response/ G.L. Patricelli, J.A.C. Uy, G. Walsh, G. Borgia// Nature. — 2002. — V.415. — P.279—280.
Patterson I.J. Aggressive interactions in flocks of rooks Corvus frugilegus L.: a study in behaviour ecology //Function and Evolution in Behaviour. – Clarendon Press Oxford, 1975. – P. 169—183.
Patterson I.J. Territorial behaviour and the limitation of population density / I.J. Patterson // Ardea. – 1980. – V.68. – P. 53–62.
Pöysä H. Feeding-vigilance trade-off in the teal (Anas crecca): effects of feeding method and predation risk/ H. Pöysä // Behaviour — 1987. — V.103. — P.108—1
Свежие комментарии