 Print
Print  PDF
PDF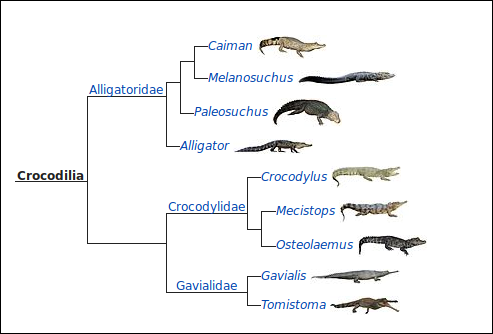
Кровеносная система крокодилов является самой сложной в животном мире. Однако, многие ее особенности недопонимаются обывателями, и благодаря стараниям консерваторов даже подвергаются неправильным интерпретациям.
Для того чтобы понять как работает сердце у крокодилов, нужно понимать общие черты строения кровеносной системы у позвоночных. Первично-водные позвоночные (традиционные классы круглоротых, хрящевых и костных рыб, кроме рыб, имеющих легочное дыхание) обходятся одним кругом кровообращения, начинающимся от двухкамерного сердца, состоящего из одного желудочка и одного предсердия. Кровь движется от сердца к жабрам, где насыщается кислородом, после чего снабжает им органы-потребители и возвращается к сердцу. Однако, наземные позвоночные существуют в более суровой сухопутной среде и как правило имеют более высокий уровень обмена веществ, чем рыбы, что стало причиной перестройки их кровеносной системы. Легкие — основной орган дыхания наземных позвоночных (кроме некоторых земноводных), намного эффективнее жабр могут насыщать кровь кислородом, но они попросту не выдержат воздействие того давления, которое нужно для того чтобы прокачать кровь через все тело. Именно поэтому в организме всех современных амниот — млекопитающих и завропсид (т.е. рептилий, включая птиц), кровь движется по двум кругам кровообращения — большому и малому. В малом (или лёгочном), кровь обогащается кислородом и избавляется от углекислого газа, проходя сквозь лёгкие, а в большом (или системном), питает кислородом все органы тела. Ни правый, ни левый круги кровообращения полноценными кругами не являются, так как замыкаются друг на друге: в малом круге кровообращения из лёгких кровь идет в левое предсердие, где выбрасывается левым желудочком в большой круг кровообращения, в то время как кровь из большого круга возвращается в правое предсердие, после чего правый желудочек вовлекает ее в малый круг кровообращения. Правая (легочная) половина сердца, в которой начинается малый круг кровообращения, при этом слабее левой, так как в ней поддерживается относительно низкое давление. Чтобы не было вопросов по терминологии, я также упомяну что венами называются те кровеносные сосуды, которые несут кровь к сердцу, а артериями — кровеносные сосуды, несущие кровь от сердца. Не полностью разделенные межжелудочковой перегородкой сердца неархозавровых рептилий (то есть, всех современных рептилий, кроме крокодилов и птиц) позволяют им смешивать насыщенную кислородом кровь, поступающую по венам малого круга кровообращения, с дезоксигенированной кровью (т.е. «венозной кровью», кровью, гемоглобин которой отдал кислород), поступающей по венам большого круга кровообращения.
Когда происходит смешивание, переход потока крови с одной стороны на другую называется «шунтированием». При этом можно наблюдать как дезоксигенированная кровь из вен системного круга кровообращения шунтируется обратно к телу в артериях большого круга кровообращения. Направленная на смешивания кровь при этом движется от правой стороны сердца к левой — это так называемый шунте справа налево. Шунт слева-направо же происходят тогда, когда некоторая часть насыщенной кислородом крови из легких возвращается обратно в малый круг кровообращения. Причины, способы контроля и корреляции шунтирования интенсивно изучались у рептилий, особенно его связь с нырянием и дыханием. Общий вывод таков, что в действительности существует широкий спектр механизмов шунтирования как у разных видов рептилий, так и при разных физиологических состояниях конкретных животных.
Также как существует обилие морфологических форм традиционно «трехкамерных» сердец. Как правило у рептилий происходит мало шунтирования при нормальном дыхании, но шунтирование в обход малого круга кровообращения выражено во время добровольного погружения и особенно во время принудительного погружения под воду в лабораторных условиях. Шунтирование имеет свои важные функции для организма рептилий, и не является какой-либо «анатомической недоработкой» (что часто пытаются сказать консерваторы, стремящиеся превратить эволюционное древо наземных позвоночных в лестницу от субъективно «менее развитого» к субъективно «более развитому»).
По мнению разных авторов оно может участвовать в поведенческой терморегуляция, перфузии кишечника после приема пищи, кожном газообмене и легочной фильтрации. Для крокодилов было показано, что шунтирование важно не только при погружении, но и позволяет снабжать органы пищеварительной системы животного кровью, богатой углекислым газом, где последний используется при выработке желудочного сока. А также то что шунтирование может играть определенную роль в утилизации молочной кислоты, являющейся продуктом анаэробного метаболизма.
В чем же заключается усложнение и уникальность кровеносной системы крокодилов? Несмотря на их четырехкамерные сердца с полностью разделенными желудочками, у крокодилов может происходить шунтирование в обход легких. Но в отличие от неархозавровых рептилии, у которых шунтирование происходит в желудочке/не разделенных желудочках, у крокодилов шунтирование идет за счет уникального расположения системных артерий и дополнительных клапанов в сердце. В отличии от млекопитающих и птиц, крокодилы, подобно неархозавровым рептилиям имеют левую и правую дуги аорты — артерии, отходящее от сердца в большом круге кровообращения.
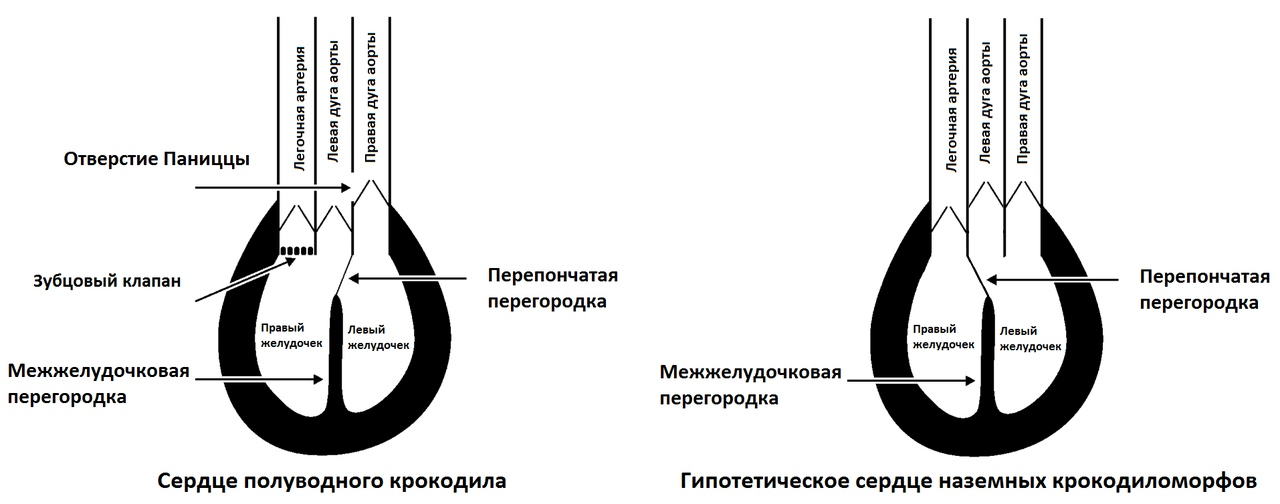
Левая дуга аорты отходит от правого желудочка (т.е. оттуда же, откуда начинается малый круг кровообращения!), а правая дуга аорты отходит из левого желудочка. Отверстие Паниццы соединяет правую и левую дуги аорты (они прилегают друг к другу) над их двустворчатыми клапанами. Наконец, правая и левая дуги аорты у крокодилов соединяются сосудом-анастомозом сосудом сзади от сердца. Легочная артерия (с которой начинается малый круг кровообращения) у крокодилов берет начало в правом желудочке, также как и у других позвоночных. Однако, малый круг кровообращения у крокодилов ограничен не одним, а двумя клапанами.
Первый из них, так называемый зубцовый клапан, является уникальным анатомическим приобретением крокодилов и может ограничивать поток. Второй клапан — это так называемый створчатый клапан, который предотвращает обратный поток крови также как и у других позвоночных. В нормальном режиме работы сердца крокодила кровь из правого желудочка поступает только в легочную артерию, в то время как доступ крови к левой дуге аорты остается закрытым за счет клапана, потому что давление в левой дуге аорты всегда больше, чем в правом желудочке. Кровь из левого желудочка поступает в правую дугу аорты, причем некоторая ее часть может пройти отверстие Паниццы в левую дугу аорты, что особенно выражено во время диастолы. Кровь также может перемещаться между обеими дугами аорты через анастомоз позади сердца.
Независимо от перемещения крови по дугам аорты, в нормальном режиме работы сердца у крокодилов происходит полное разделение потоков оксигенированной (богатой кислородом «артериальной») и дезоксигенированной (бедной кислородом «венозной») крови, также как у млекопитающих и птиц. Однако, в определенных условиях сжатие зубцового клапана перекрывает малый круг кровообращения и приводит к тому, что давление в правом желудочке превышает давление в левой дуге аорты, и двустворчатый клапан левой дуги аорты открывается.
Дезоксигенированная кровь поступает в большой круг кровообращения и происходит шунтирование. Кровь, выброшенная в левую дугу аорты, идет главным образом к органом пищеварительной системы, но есть свидетельства того, что некоторая ее часть может пойти в обратном направлении через отверстие Паниццы в правую дугу аорты.
Проксимальная (ближняя) часть левой дуги аорты расширена, в отличие от ее более дистальных (отделенных) частей, предполагая, что через нее может проходить довольно большой объем крови из правого желудочка. Шунтирование, безусловно, контролируется самими крокодилами, и инициируется сужением зубцового клапана у основания малого круга кровообращения. Зубцовый клапан работает под нервным управлением.
Адренергическая стимуляция ослабляет зубцовый клапан, вероятно, оставляя его открытым во время условия стресса или физических упражнений. Размер отверстие Паниццы также контролируется, адренергическая стимуляция уменьшает его диаметр, тогда как вазоактивный кишечный полипептид увеличивает его.
И главное вишенкой на торте является то, что размещение левой дуги аорты в правом желудочке, зубцовый клапан и отверстие Паниццы, безусловно, являются уникальными приобретениями крокодилов, развившимися при переходе их наземных и почти наверняка теплокровных предков к полуводному образу жизни. Основной функцией четырехкамерного сердца, по всей видимости, является разделение системного и легочного кровяного давления. Это разделение необходимо не только для достижения высокого уровня метаболизма, но и для транспорта крови на большие расстояние. В обоих случаях необходимо высокое давление в системном круге кровообращения, и низкое — в легочном. При этом даже анатомически неразделенные желудочки сердец некоторых неархозавровых рептилий (например, варанов, которые ведут активный образ жизни, а также крупных наземных змей, которые достигают большой длины) могут создавать заметное разделение давления легочного и системного кругов кровообращения, что отражает возможность его достижения без развития четырехкамерного сердца.
У неархозавровых рептилий разделение давления легочного и системного кругов кровообращения достигается за счет разделения желудочков мускульными выростами, происходящее во время систолы. Несмотря на четырехкамерное сердце и эффективное разделение системных и легочных давлений, среднее кровяное давление в большом круге кровообращения у крокодилов находятся в пределах такового у холоднокровных животных (значения в 3,9-5,3 кПа были получены для миссисипского аллигатора и 6,2-7,9 кПа для гребнистого крокодила). Развитие четырехкамерного сердца казалось бы нелогичным для холоднокровных животных, но отлично согласуется с тем, что предки крокодилов были теплокровными животными, требовавшими постоянно высокое кровяное давление (при этом следует также учитывать небольшой размер древнейших известных крокодиломорфов). Хотя эмбриогенез далеко не всегда повторяет филогенез (см. критику закона Геккеля), детали эмбрионального развития крокодилов также подтверждают эту гипотезу. В первую очередь — показано, что отверстие Паниццы и и зубцовый клапан у крокодилов развиваются после развития полноценного 4-камерного сердца. Детали развития перепончатой перегородки (части межжелудочковой перегородки), разделяющий левый и правый желудочки сердца, отличаются у крокодилов, птиц и млекопитающих, в связи с различным состоянием дуг аорты у этих животных.
У крокодилов перепончатая перегородка развивается и сохраняется при завершении эмбрионального развития между левой и правой дугами аорты. У птиц она развивается между легочной артерией и левой дугой аорты, но последняя быстро атрофируется и остается только правая дуга аорты. Это важное доказательство того, что птичье сердце в процессе эволюции не проходило через стадию сердца крокодилов с левой дугой аортой, открывающейся в правый желудочек — это две эволюционно-независимые друг от друга конструкции. Однако, подобная особенность достаточно часто возникает у птичьих эмбрионов в качестве аномалии, чтобы быть замеченной эмбриологами. Сходство между сердцами птиц и нептичьих рептилий позволяют легко визуализировать эволюцию от общего предка, объяснив ее простым сдвигом перепончатой перегородки. Очень вероятно что сдвиг перепончатой перегородки с ее размещением между левыми и правыми дугами аорты произошел у крокодилов при переходе к полуводному образу жизни и необходимостью возникновения контролируемого шунтирования, в то время как у наземных предков крокодилов из числа крокодиломорфов, также как и у птиц, перепончатая перегородка закладывалась между легочной артерией и левой дугой аорты.
У млекопитающих неправильное развитие перепончатой перегородки с ее присоединением в неправильном месте создает наиболее распространенные врожденные дефекты перегородки, включая тетраду Фалло и полную транспозицию легочной артерии и аорты. Люди при нарушениях в развитии сердца могут даже сохранить обе дуги аорты (в норме в процессе эмбрионального развития у млекопитающих редуцируется правая) и жить с ними при условии, что через них проходит достаточное количество крови. Эти врожденные аномалии могут имеют многофакторную генетическую основу. Распространенность таких дефектов говорит о том, что лишь незначительные сдвиги в развитии перепончатой перегородки могут привести к большим различиям в окончательной морфологии сердца, при том что животное по прежнему может оставаться жизнеспособным.
За большими подробностями рекомендую к прочтению статью «Evidence for Endothermic Ancestors of Crocodiles at the Stem of Archosaur Evolution«, написанную Р. С. Сеймуром и соавторами еще в 2004 году. А также статью «Quirky cardiology: Crocs’ hearts may aid their digestion» от B. Harder (2006) о дополнительных функциях шунтирования у крокодилов.
На рисунке — схематичное изображение сердце полуводного крокодила без вен и предсердий (слева) и гипотетического сердца теплокровного наземного предка крокодилов (справа) на основе Figure 5 из Seymour et al. (2011).
Свежие комментарии